

서 론
재료 및 방법
공시재료와 형태적 특성 분석
통계분석
결과 및 고찰
한국, 일본 및 중국에서 수집한 기장 계통들의 형태적 변이
한국, 일본 및 중국에서 수집된 기장 계통들에 대한 주성분 분석
요 약
서 론
기장(Panicum miliaceum L.)은 외떡잎식물 벼목 화본과의 한해살이풀로 온대성 식물로 잡곡류에 속한다. 신석기시대부터 중앙아시아, 중동지역, 중국 등의 지역에서 재배되어 온 것으로 알려졌고, 현재도 네팔, 러시아, 미국, 스리랑카, 인도, 중국, 파키스탄 등의 온대 기후에서 널리 재배되고 있다. 주로 동아시아의 국가들은 식량작물로 이용하였고, 유럽 및 미국에서는 사료용으로 이용하였다(Graybosh and Baltensperger, 2009; Kim et al., 2013; Oelke et al., 1990; Salini et al., 2010). 한국에서 주로 재배되고 있는 기장은 영문 명이 Common millet (Panicum miliaceum L.)로 주로 중국, 일본, 인도 등에서 많이 재배되고 있다. 타 작물과 비교하여 요수량이 낮아 가뭄 적응성이 높고, 재배기간이 짧으면서 보통 식물이 자라기 힘든 개간지나 척박한 환경에도 잘 적응하며 출수 생태 및 재배 시기에 따라 농업적 특성 변이가 다양하게 나타나고 있다. 예전부터 구황작물로 재배되어 왔으며, 최근 봄, 여름 모두 재배가 가능하고 재배기간이 짧아 다양한 작부체계 활용 증가로 재배면적이 늘어나고 있다(Jeju Agricultural Research and Extension Services, 2014; Park et al., 1999). 종실의 배유 특성에 따라 찰기장과 메기장으로 나뉘고, 4월말에 파종하여 7월중에 조기 수확이 가능한 기장과 5월에서 7월중 파종하여 10월경에 수확이 가능한 기장 등 재배 시기에 따라 여러 생태형으로 나눌 수 있다(Park et al., 2015). 한국 및 중국, 일본, 인도에서 많이 재배되는 기장은 크기가 작은 다른 잡곡류와 비교하여 단백질 및 무기성분 함량이 다소 높다고 보고되고 있다(Park et al., 1999). 또한 기장은 cholesterol과 phytate를 낮추고, 특정 암을 억제하며, 심장과 간 질환을 예방하고, 간 기능 장애와 당뇨병을 관리하는 의학적 이점이 있다고 보고되었다(Saha et al., 2016; Zhang et al., 2014). 쌀, 밀, 옥수수 등과 동일한 무게와 양을 기준으로 비교하였을 때 알곡이 작아서 높은 영양 성분을 많이 함유하는 배아 및 호분층의 비율이 높으며, 다른 곡물류에 비하여 미네랄, 식이섬유, 비타민A 및 지방 등의 함량이 높은 영양학적 특징이 있다고 한다(Amadou et al., 2013; Ha and Lee, 2001; Oelke et al., 1990). 이러한 중요성에도 불구하고 한국에서 기장 품종은 지역 재래종을 중심으로 재배되었고, 재래종은 종자의 혼입도가 높고 수량성이 낮아 순도가 높고 생산량이 높은 기장을 생산하기가 힘들다. 그러므로 수집한 재래종들에 대하여 순계 분리 및 교잡 육종을 통하여 수량성이 높고 기능성 물질을 다량 함유하고 있는 우수한 기장 품종의 육성이 앞으로 필요하다(Ko et al., 2018).
한편 오늘날 지구상의 많은 식물 종들은 19세기 이후 산업이 발전하면서 급진적인 산업화와 도시화가 이루어지고, 국토개발과 환경파괴로 인해 식물 종의 유전적 다양성이 사라지고 있다. 더욱이 육종기술의 발달에 따른 신품종의 개발과 보급에 의하여 농가에서 예전부터 재배되고 있었던 재래종들의 유전적 침식을 가속화 시켰다. 이러한 상황을 개선시키기 위해서는 다양한 유전자원의 지속적인 탐색과 수집을 통해 보전하는 것이 중요하다. 잡곡 중 하나인 기장은 재배면적이 1970년대 약 1,000 ha 정도로 알려졌으나, 계속 감소되어 현재 산간지 및 해안지 등에서 특수 가공 목적으로 재배되고 있다(Yoon et al., 2008). 이처럼 경제성이 낮은 작물로 알려진 잡곡은 재배면적이 점차 감소하고 있었지만, 최근 잡곡에 대한 기능성과 영양성이 다시 알려지면서, 잡곡의 소비와 선호도가 점차 늘고 있으며, 2007년 이후 69,000톤으로 수요량이 크게 증가하였다. 그러나 수요량 중 약 50,000톤은 수입에 의존하고 있는 상황이다(MIFAFF, 2010). 또한 잡곡은 천연색소 및 식·의약품 등의 소재로 다양한 용도로 사용이 가능하며, 관상 및 환경보전 작물로도 우수하여 새로운 부가 가치를 만들어 낼 수 있는 작물로 여러 연구가 활발히 진행 중에 있다(Jung et al., 2016).
오늘날 작물의 육종연구는 다양한 종류의 유전자원을 수집하여 평가함으로써 유전 및 육종 연구의 중요한 재료로 이용하고 있다. 따라서 유전자원을 수집하고 이들의 형태 및 유전적 특성을 활용한 우수한 유전자원 선발은 작물의 육종 효율을 향상시키기 위해 매우 중요하다. 본 연구는 기장의 유전자원의 다양성 확보와 신품종 개발을 위해 한국, 일본 및 중국에서 수집하여 농촌진흥청 국립농업과학원 농업유전자원센터에서 보존하고 있는 75계통의 기장 자원들에 대하여 형태적 특성을 이용한 주성분 분석(PCA 분석)을 실시하였다. 본 연구 결과는 앞으로 국내 환경에 적합한 우수한 기장 신품종 개발을 위한 유전‧육종 연구에 유용한 정보를 제공할 것으로 기대한다.
재료 및 방법
공시재료와 형태적 특성 분석
한국, 일본 및 중국에서 수집한 기장 유전자원들에 대한 형태적 특성 및 변이성을 조사하기 위해 한국(54계통), 일본(12계통), 중국(9계통)에서 수집한 총 75계통을 농촌진흥청 국립농업과학원 농업유전자원센터에서 분양 받았고, 이들 자원에 대한 기본 정보는 Table 1에 제시하였다.
Table 1.
List of 75 common millet accessions of RDA-Genebank used in this study
형태적 특성을 조사하기 위해 계통 당 30립씩 종자를 파종하여 2019년 춘천시 천전리에 위치한 강원대학교 부속농장에서 30립 중 발아한 20개체를 1주 2본씩 이식하였다. 기장 개체 간 거리를 25 cm, 기장 계통 간 거리를 75 cm 간격으로 흑색 P. E. 필름으로 멀칭한 포장에서 일반 관행 재배법 준하여 재배하였다. 형태적 특성 조사는 각 계통 별로 10개체씩 조사하였으며, 형태적 조사 형질은 종피색(QL1), 수형(QL2), 간장(QN1), 개화일수(QN2) 등 비교적 식별이 편리한 양적 및 질적 형질들에 대하여 생육 중기부터 수확기까지 생육시기별로 조사하였다(Table 2).
Table 2.
Characteristics used in the morphological analysis for 75 common millet accessions
통계분석
기장의 계통 별 형태적 변이와 다양성을 분석하기 위하여 한국 및 일본과 중국에서 수집한 계통들에 대한 형질 별 변이를 비교 분석하였다. 주성분 분석(Principal Component Analysis)은 각 집단 간 그리고 집단 내의 계통들 사이에서의 형태적 변이성을 알아보기 위하여 수행하였다. 본 실험에서 수행한 주성분 분석은 Microsoft Excel 통계프로그램을 이용하여 분석하였다.
결과 및 고찰
한국, 일본 및 중국에서 수집한 기장 계통들의 형태적 변이
지구상에 재배되고 있는 작물 재래종들의 형태적 및 생태적 변이는 서로 다른 환경에서의 작은 진화적 변화(microevolutionary changes)에 의해서 일어나고 있다(Gould and Johnston, 1972; Wyatt and Antonovics, 1981; ). 따라서 자연적으로 폭 넓게 분포하고 있는 어떤 재래종 작물의 지리적 분포에 따른 형태적 및 생태적 변이는 그 작물의 진화 및 분화과정을 연구하는데 있어 매우 유용한 정보를 제공하게 된다(Schwanitz, 1966; Harlan, 1992). 본 연구는 한국을 포함하여 일본, 중국에서 수집한 재래종 기장 75계통들에 대하여 지리적 분포에 따른 형태적 변이를 조사하였다. 본 연구에서는 한국, 일본과 중국에서 수집된 기장 75계통(한국 54계통, 일본 12계통, 중국 9계통)을 이용하여 2개의 양적 형질과 2개의 질적 형질에 대한 특성조사를 실시한 결과는 다음과 같다(Table 3).
Table 3.
Mean, standard deviation, range and accession number for 2 quantitative and 2 qualitative characters among 75 common millet accessions
1) 간장(QN1) – 한국 기장 계통들의 간장은 평균 143.4 ± 9.5 cm로, 최소 115.2 cm부터 최대 166.4 cm 값의 범위였으며, 일본 기장 계통들은 평균 140.1 ± 19.6 cm로, 86.3 cm부터 158.4 cm 값의 범위를 보였다. 반면에 중국
기장 계통들은 평균 95.3 ± 15.8 cm로 75.8 cm부터 121.7 cm의 값을 나타내었다.
2) 출수일수(QN2) – 한국 기장 계통들의 출수일수는 평균 87.6 ± 4.7일로 최소 69일부터 최대 95일까지의 범위를 나타내었다. 일본 기장 계통들의 평균 85.0 ± 7.9일로 69일부터 95일까지의 범위를 나타냈고, 중국에서 수집한 계통들은 평균 53.0 ± 3.0일로 49일부터 55일의 범위를 나타내었다.
3) 종피색(QL1) – 한국 기장 계통들의 경우 갈색(8계통), 백색(21계통), 암갈색(10계통), 줄무늬암갈색(3계통), 황백색(11계통), 회갈색(1계통) 등으로 나타났고, 일본 기장 계통들은 갈색(6계통), 백색(3계통), 암갈색(1계통), 황백색(2계통) 등으로 나타났다. 중국 기장 계통들은 갈색(3계통), 백색(2계통), 암갈색(4계통) 등으로 나타났다.
4) 수형(QL2) – 한국 기장 계통들의 경우 산수형(4계통), 편밀수형(3계통), 편산수형(12계통), 편수형(35계통) 등으로 나타났고, 일본 기장 계통들은 편밀수형(8계통), 편수형(4계통)으로 나타났다. 중국 기장 계통들은 밀수형(3계통), 산수형(3계통), 편밀수형(1계통), 편수형(2계통) 등으로 나타났다.
본 연구에서 각 나라별로 수집한 기장 자원들에 대한 형태적 특성조사에서 일부 형질을 제외하고 대부분의 형질들은 국가 간 비슷한 특성을 나타내었다. 간장의 경우 한국과 일본에서 수집한 계통들은 비슷한 간장을 나타내고 있으나, 중국에서 수집한 계통들은 한국과 일본의 계통들보다 비교적 짧은 간장을 나타내고 있었다. 이것은 수집한 지역의 기후 및 계통 별 생육기간인 출수일수와 직접적으로 관련이 있는 것으로 판단되는데, 한국과 일본에서 수집한 계통들은 비슷한 출수일수를 나타내고 있으나 중국에서 수집한 계통들은 한국과 일본의 계통들보다 빨리 개화하는 조숙성 계통들로만 구성되어 있었다. 이러한 결과는 아마도 중국지역에서 수집한 계통들의 경우 북쪽지방에서 수집한 된 것으로 생각되었다. 이전의 Lee and Ohnishi (2001); Ma and Lee (2017) 등의 보고에 의하면 들깨 작물의 경우 중국 남부에는 대부분이 만생형 계통들이 재배되고 있으나, 북부지역에는 조‧중생형 계통들이 재배되고 있는 것으로 보고하였다. 한편 Yoon et al. (2008)은 한국의 기장 계통들에 대한 특성조사에서 평균 간장이 181.4 cm, 평균 출수일수가 73.7일로 보고하였는데, 본 연구에서도 비슷한 경향을 나타내고 있었다. 본 연구결과는 한국을 포함한, 일본과 중국의 재래종 기장 계통들의 지리적 분포에 따른 형태적 변이를 이해하는데 유용한 정보를 제공할 것으로 기대한다.
한국, 일본 및 중국에서 수집된 기장 계통들에 대한 주성분 분석
한국, 일본과 중국에서 수집한 기장 계통들에 대한 지리적 분포에 따른 형태적 변이를 확인하기 위해서 기장 75계통에 대하여 2개의 양적 형질과 2개의 질적 형질을 이용하여 주성분 분석(PCA 분석)을 실시하였다. 그 결과, 1차 및 2차 component는 전체 변이의 52.2%와 24.4%를 나타내어 전체 76.6%의 변이를 나타냈다(Table 4). 조사된 형질들 중에서 종피색(QL1), 수형(QL2), 간장(QN1), 출수일수(QN2)는 제 1주성분에서 모두 양의 방향으로 기여하였고, 제 2주성분에서 종피색(QL1)은 양의 방향으로 기여하였고, 수형(QL2), 간장(QN1), 출수일수(QN2)은 음의 방향에 기여한 것으로 나타났다.
Table 4.
Eigen vectors and cumulative variance of the first and second principal components
기장 75계통들에 대하여 주성분 분석 결과를 이용하여 scatter diagram 분석을 실시하였다(Fig. 1). 그 결과 제 1주성분(PC1)과 제 2주성분(PC2)을 기준으로 4개의 사분면으로 구분되었다. 한국 기장 계통들은 대부분이 제 1사분면과 제 4사분면에 분포하고 있으나, 일부 계통(IT 109200, IT 185528, IT 103775, IT 185540)들은 제 2사분면과 제 3사분면에 각각 분포하였다. 중국 기장 계통들은 모두 제 2사분면과 제 3사분면에 분포하였으며, 일본 계통들은 주로 제 3사분면과 제 4사분면에 분포하였지만 Japan 9 (IT 297394) 계통은 2사분면에, Japan 1 (IT 297380) 계통과 Japan 10 (IT 297395) 계통은 제 1사분면에 각각 분포하였다.
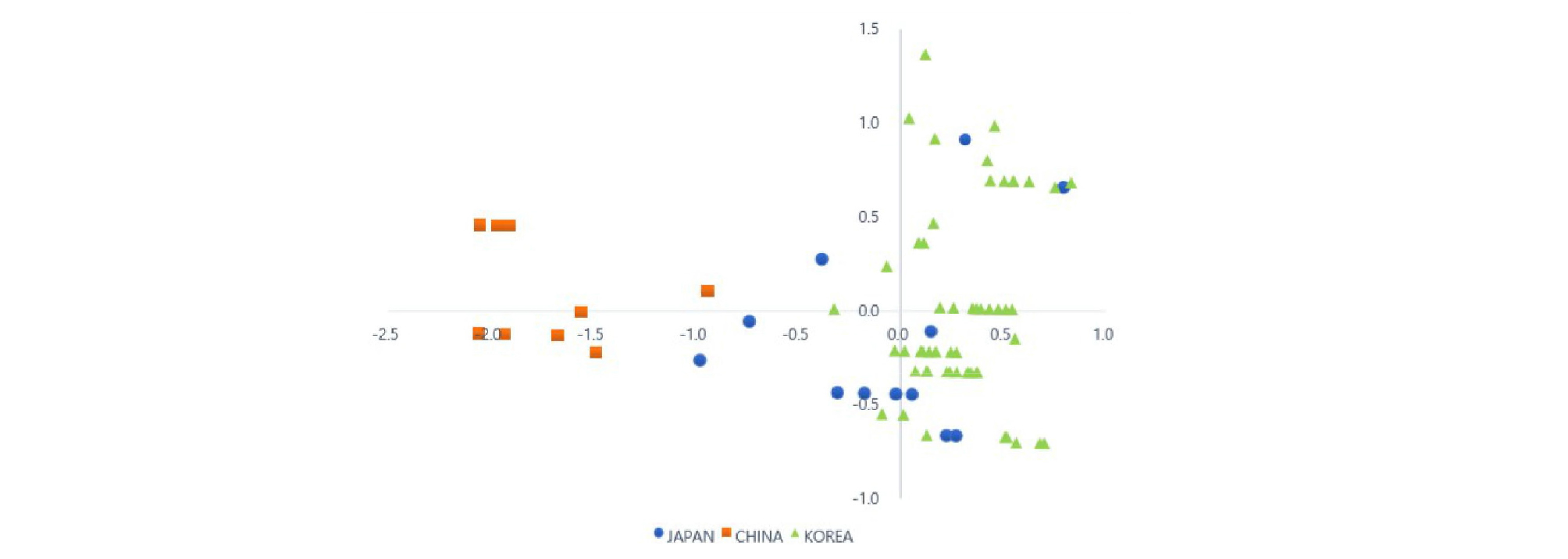
이상의 결과에서 한국, 일본과 중국의 국가별로 기장 계통들 사이의 구분이 명확하지 않아 종피색, 수형, 출수일수 등으로 나누어 분석해보았다. 그 결과 출수일수의 경우 사분면에서 구분이 명확하지 않았으나, 수형을 기준으로 한 분석에서는 밀수형 계통들이 3사분면에 위치하였고, 종피색을 기준으로 분석에서는 황백색과 줄무늬암갈색 계통들이 1사분면에 위치하였다. 본 연구에서 한국, 일본 및 중국의 75계통들은 갈색(Brown), 백색(White), 암갈색(Dark brown)의 종피색을 가진 계통이 모두 존재하므로 국가별 계통 특성을 명확히 구분하기가 어려웠다. 이삭의 수형에서는 한국 기장 계통들은 대부분 편수형(Contractum) 계통이 많이 있는 것으로 확인되었고, 일본은 편밀수형 계통들이 많이 있는 것으로 확인되었으나, 밀수형(Compactum) 계통들은 모두 중국 계통들에서만 나타났다. 본 연구에서 수행한 주성분 분석 결과에서 종피색과 수형을 기준으로 동아시아에서 수집한 기장 계통들을 명확히 구분하기는 어려운 것으로 생각되었다. 이러한 결과는 Cho et al. (2010)의 보고와 같이 동아시아 지역의 비슷한 기후 환경 내에서 자연적 선택 또는 인간에 의한 종자 이동 등에 의해 발생한 것으로 생각되었다.
이상의 결과에 의하면 주성분 분석에 의해 동아시아에서 수집한 75계통의 기장을 지리적으로 명확하게 식별할 수는 없었다. 본 연구에서 분석에 이용한 중국 기장 계통들은 생육기간이 짧고 출수가 빠르며, 간장이 한국과 일본 계통들에 비해 크지 않아 구분이 가능했으나, 한국과 일본에서 수집한 기장 계통들은 서로 명확히 구분되지가 않았는데 이것은 이들 지역에서 수집한 기장 계통들이 지리적 분포와 관련이 있는 것으로 생각되었다. 따라서 본 연구결과는 주성분 분석을 통해 동아시아에서 수집한 재래종 기장 계통들의 형태적 변이를 이해하는데 유용한 정보를 제공할 것으로 기대한다.
요 약
본 연구는 한국을 포함하여 일본, 중국에서 수집한 재래종 기장 75계통들에 대하여 지리적 분포에 따른 형태적 변이를 조사하였다. 본 연구에서는 한국, 일본과 중국에서 수집된 기장 75계통(한국 54계통, 일본 12계통, 중국 9계통)들에 대하여 2개의 양적 형질과 2개의 질적 형질을 이용하여 형태적 특성을 조사하였다. 그 결과 각 나라별로 수집한 기장 자원들은 일부 형질을 제외하고 대부분의 형질들은 국가 간 비슷한 특징을 나타내었다. 간장의 경우 한국과 일본에서 수집한 계통들은 비슷한 크기의 간장을 나타내고 있으나 중국에서 수집한 계통들은 짧은 간장을 나타내고 있었다. 주성분 분석(PCA 분석)에서 1차 및 2차 component는 전체 변이의 52.2%와 24.4%를 나타내어 전체 76.6%의 변이를 나타냈다. 조사된 형질들 중에서 종피색(QL1), 수형(QL2), 간장(QN1), 출수일수(QN2)는 제 1주성분에서 양의 방향으로 기여하였고, 제 2주성분에서 종피색(QL1)은 양의 방향으로 기여하였고, 수형(QL2), 간장(QN1), 출수일수(QN2)은 음의 방향에 기여한 것으로 나타났다. 주성분 분석 결과에서 한국, 일본과 중국의 기장 계통들 사이를 명확히 구분하지 못하였다. 본 연구결과는 한국을 포함한 일본과 중국의 재래종 기장 계통들의 지리적 분포에 따른 형태적 변이를 이해하는데 유용한 정보를 제공할 것으로 기대한다.
