

서 론
재료 및 방법
반수체 유기체의 국내 적응성 검정
활용 집단의 반수체 유기율 조사
염색체 배가 시기 비교
배가 반수체 기술에 의해 육성된 계통의 농업형질 평가
통계분석
결과 및 고찰
반수체 유기체의 국내 적응성 검정
활용 집단의 반수체 유기율 조사
염색체 배가 시기 비교
배가 반수체 기술에 의해 육성된 계통의 농업형질 평가
결 론
서 론
옥수수 품종육성은 계통육성과 육성된 계통을 이용한 교잡종의 생산력검정을 통한 우량 교잡종 선발의 단계로 이루어진다. 이 단계 중에서 계통 육성은 99% 이상의 순도를 가지는 순계를 육성하는 과정이다. 옥수수는 타가수정 작물이므로 계통육성을 위해 7세대 이상 인공교배를 통하여 순도를 높이고 선발하는 과정이 필요하다. 국내의 옥수수 육종은 이러한 전통적인 방법에 전적으로 의존하고 있다.
계통육성에서의 이러한 어려움을 극복하고자 쉽고 빠르게 계통을 육성할 수 있는 신기술인 배가 반수체(Doubled Haploid) 기술이 개발되어 이용되고 있다. 배가 반수체 기술은 정상 식물체(2n)에서 반수체(n)를 유기하고, 이를 배가시켜 순도 100%의 옥수수 계통을 육성하는 기술이다(Wu et al., 2020). 배가 반수체 기술을 이용한 옥수수 계통육성은 다음과 같은 과정을 통하여 3세대 이내에 100% 동형접합체를 만들 수 있다. ①옥수수 집단을 모본으로 하고 반수체를 유기하는 유기체(inducer)를 부본으로 하여 교배를 통한 반수체(n) 유기, ②색소 유전자인 R1-nj의 유무에 따른 반수체 종자 선별, ③선별 반수체 종자를 발아시킨 옥수수 유묘 단계에서 염색체 배가, ④그리고 인공교배를 통한 계통 생산(Chang and Coe, 2009).
전통적인 육종방법과 비교할 때 배가 반수체 기술을 이용한 옥수수 계통육성은 육종연한을 7세대 이상에서 3세대 이내로 크게 단축시킬 수 있다. 또한 시험포장 규모를 축소할 수 있으므로 계통 육종에 필요한 시간, 노동력 및 예산을 절감할 수 있다(Ren et al., 2017). 배가 반수체 기술을 통하여 육성된 계통은 이론상으로 순도 100%의 순계이다. 이는 99.9%의 순도는 가능하지만 100%의 완전한 동형접합체 육성은 불가한 전통육종방법과 대비되는 배가 반수체 육종방법의 특징이다(Chaikam et al., 2019).
단기간에 대량의 계통 육성이 가능하므로 배가 반수체 기술을 이용하면 유전자지도 작성 및 분석을 위한 집단 양성이 용이하다. 또한 옥수수 계통육종 과정에서 목표형질의 선발을 위해서 필요한 식물체의 개체 수를 4n에서 2n으로 획기적으로 감소시킬 수 있다(Prasanna et al., 2012). 배가 반수체 기술을 분자마커와 연계하여 이용할 경우 marker-assisted selection (MAS), 유전자 도입, 유전자 집적 등에도 유용하게 이용될 수 있다. 또한 선진국에서는 자식계통의 수가 증가함에 따라 계통의 효율적 선발을 위한 연구가 대두되었으며, 분자마커를 활용하여 계통선발의 효율을 높이고 있다. 이러한 배가 반수체 기술의 다양한 장점 때문에 글로벌 종자회사 대부분은 계통육종의 대부분을 배가반수체 기술에 의존하고 있다. 그러나 국내 옥수수연구에서 배가 반수체 기술을 이용하는 곳은 강원도농업기술원 옥수수연구소가 유일하며, 배가 반수체 기술을 적용하는 초기단계라고 할 수 있다(Lee et al., 2014).
본 연구는 옥수수 연구분야에서 국내에 적용된 바가 없는 배가 반수체 기술을 도입하고 정착시키고자 수행하였다. 그 결과로서 반수체 유기체의 국내 적응성, 옥수수 집단과의 교배를 통한 반수체 유기율 평가, 반수체 종자의 염색체 배가 시기 비교, 그리고 배가 반수체 기술에 의해 육성된 계통의 농업형질에 대하여 보고하고자 한다.
재료 및 방법
반수체 유기체의 국내 적응성 검정
강원도농업기술원 옥수수연구소는 국제옥수수·밀연구소(CIMMYT) 및 독일의 Hohenheim 대학과 계약을 통하여 반수체 유기체인 TAILs를 상업적으로 이용할 수 있는 권리를 확보하였고 이를 본 연구에 활용하였다. 확보한 반수체 유기체는 국제옥수수·밀연구소(CIMMYT)에서 열대형과 온대형을 교배하여 아열대형으로 개량한 유기체이다. 아열대형 반수체 유기체인 TAILs (Prasanna et al., 2012; Prigge et al., 2012)의 국내 적응성 검정을 위하여 2014년 4월 23일 육묘포트에 파종하여 관리하였고, 5월 14일 옥수수연구소 연구포장에 정식하였다. 무기질 비료의 시용량은 N-P2O5-K2O로 10 a당 17.4-3-6.9 kg이었고, 퇴비는 1.5톤을 시용하였다. 질소질 비료의 반량은 밑거름으로, 반량은 7-8엽기에 웃거름으로 주었으며, 인산질 비료와 칼리질 비료 및 퇴비는 모두 밑거름으로 시용하였다.
활용 집단의 반수체 유기율 조사
옥수수 집단으로부터 반수체를 유기하기 위하여 옥수수 집단을 모본으로 하고 반수체 유기체인 TAILs를 부본으로 하여 인공교배를 실시하였다. 2014년부터 2017년까지 찰옥수수 및 종실용옥수수 48집단을 이용하였다. Bernardo (2009)는 배가 반수체 기술을 이용하는데 가장 적합한 집단의 형태는 F2집단이라고 하였다. 이에 따라 본 연구에 활용한 집단은 대부분 F2를 이용하였으며, 일부집단은 F1을 이용하였다. 옥수수 집단은 매년 5월 초에 옥수수연구소 연구포장에 직파하였다. 반수체 유기체인 TAILs는 육묘포트에 파종하여 관리한 후 연구포장에 정식하였다. 이용한 옥수수 집단의 개화시기가 상이하기 때문에 옥수수 집단과의 교배시기를 맞추기 위하여 유기체는 집단 파종 7일 전부터 10일 간격으로 4회 파종하였다.
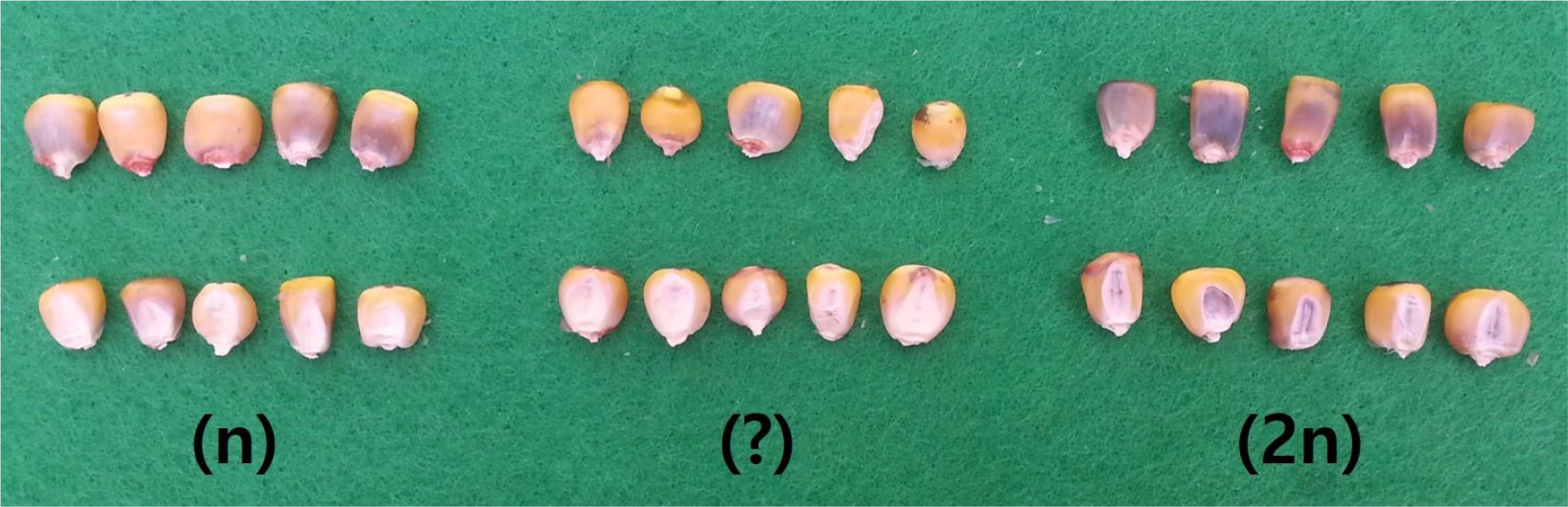
옥수수 집단과 반수체 유기체의 교배에 의해 생산된 종자는 Nanda와 Chase (1966), Prasanna 등(2012) 등이 보고한 선별방법에 따라 배와 배유의 색소 발현을 기준으로 반수체(n)와 이배체(2n)로 구분하였다(Fig. 1). 반수체 종자는 배유에는 색소 발현이 있으나 배에는 안토시아닌 색소가 발현되지 않는다. 이배체 종자는 배와 배유에 모두 자색의 안토시아닌 색소가 발현된다. 배에서의 색소 발현이 없는 종자를 반수체 종자로 선별하였으며, 또한 배에서 색소 발현 유무가 불명확하여 구분이 어려운 종자도 반수체 종자에 포함하였다.
염색체 배가 시기 비교
반수체(n)로 선별된 종자는 염색체 배가에 활용하였다. 염색체 배가는 발아종자 및 유묘 등 2가지 생육단계에서 처리하였다. 발아종자는 Prasanna 등(2012)의 방법에 따라 발아시험용 paper에 종자를 넣고 28°C에서 3일간 발아를 시킨 후 발아된 종자를 colchicine 0.04% 용액에 12시간 침지하여 배가처리 하였다. 배가처리 후 발아종자는 지피포트에 육묘하여 약 2주 이상 활착시킨 후 배가반수체 시설하우스에 정식하였다. 유묘는 Melchinger 등(2016)의 방법에 따라 육묘포트에 반수체 종자를 파종하여 약 2엽기까지 생장하였을 때 식물체를 뽑아서 뿌리의 상토를 세척한 후 유묘를 colchicine 0.07% 용액에 5시간 침지하여 배가처리 하였다. 배가처리 후 유묘는 지피포트에 육묘하여 약 7일 이상 활착시킨 후 배가반수체 시설하우스에 정식하였다.
반수체 종자의 선별은 배에서의 안토시아닌 색소발현을 육안으로 선별하는 것이므로 선별오류가 발생하기 쉽다. 따라서 정식 후 식물체가 생장함에 따라 초세가 다른 이배체(2n)는 제거하였다. 배가 처리한 식물체에서의 화분생산은 소량이므로 계통을 육성하기 위한 인공교배는 화분량이 극히 적은 경우 3회까지도 실시하였다.
배가 반수체 기술에 의해 육성된 계통의 농업형질 평가
배가 반수체 기술에 의해 육성된 계통에 대하여 예비 선발을 실시하였고, 이 중 우량한 종실용 5계통, 찰옥수수 5계통에 대한 농업형질을 평가하였다. 2017년 5월 4일 육묘포트에 파종하여 관리하였고, 5월 19일 옥수수연구소 연구포장에 난괴법 3반복으로 배치하여 정식하였다. 무기질 비료의 시용량은 N-P2O5-K2O로 10 a당 17.4-3-6.9 kg이었고, 퇴비는 1.5톤을 시용하였다. 질소질 비료의 반량은 밑거름으로, 반량은 7-8엽기에 웃거름으로 주었으며, 인산질 비료와 칼리질 비료 및 퇴비는 모두 밑거름으로 시용하였다.
통계분석
통계분석을 위해 R scripts (R Core Development Team, 2015)를 사용하였다. 연도별 반수체 유기율의 차이를 파악하기 위해 크루스칼 왈리스(Kruskal-Wallis) 검정을 실시하였고 계통간 주요 농업 형질의 차이를 판단하기 위해 분산분석을 실시하였다. 발아묘와 유묘의 반수체율 비교를 위해 독립표본 t-검정을 실시하였다.
결과 및 고찰
반수체 유기체의 국내 적응성 검정
강원도농업기술원 옥수수연구소에서 확보한 아열대형 반수체 유기체를 강원도 홍천에서 재배하였을 때의 농업형질은 Table 1과 같다. TAILs의 출사기는 7월 4일이었으며 파종에서 출사기까지의 일수는 72일이 소요되었다. 간장은 196 cm, 착수고는 83 cm로 착수고율은 42%였다. 이삭장은 14.6 cm, 100립중은 35.7 g이었으며 종피색은 진보라색을 띄었다. TAILs의 양친인 P1과 P2의 농업형질을 비교해보면, P1의 출사일수는 72일, 간장은 158 cm, 착수고는 79 cm로 착수고율은 50%, 이삭장은 9.5 cm, 100립중은 29.4g이었으며 종피색은 진보라색의 띄었다. P2의 출사일수는 79일, 간장은 187 cm, 착수고는 87 cm로 착수고율은 47%, 이삭장은 10.4 cm, 100립중은 28.8 g이었으며 종피색은 보라색의 띄었다. 출사일수 및 종피색은 TAILs와 P1이 유사하였고, 이삭장과 100립중은 P1과 P2가 유사하였다.
Table 1.
Agronomic characteristics of sub-tropical haploid inducers, TAILs, and parents in South Korea
| Inducers | Silking date |
Days to silking (days) |
Pollen shed period |
Plant height (cm) |
Ear height (cm) |
Ear length (cm) |
100 seed weight (g) | Seed color |
| TAILs1) | July 4 | 72 ± 1.82) | July 1-July 11 | 196 ± 8.4 | 83 ± 4.6 | 14.6 ± 0.7 | 35.7 ± 0.9 | Dark purple |
| P13) | July 4 | 72 ± 1.5 | July 1-July 11 | 158 ± 7.9 | 79 ± 3.5 | 9.5 ± 0.6 | 29.4 ± 0.7 | Dark purple |
| P2 | July 9 | 79 ± 1.6 | July 4-July 15 | 187 ± 8.3 | 87 ± 2.8 | 10.4 ± 0.8 | 28.8 ± 1.1 | Purple |
열대형 또는 아열대형 옥수수가 온대지역에서 생육할 경우 영양생장 기간이 길어지거나 심한 경우 생식생장으로 전환하지 못하는 경우도 발생한다. 확보한 반수체 유기체는 아열대형 자원이므로 국내에서 정상적으로 영양생장 및 생식생장을 하는지 확인하는 것이 필요하였다. TAILs는 도복에 약한 특성이 있었지만 다른 농업형질은 일반적인 옥수수 자원으로 평가되었으며, 국내에서 많이 재배하는 미백2호와 비교하면 TAILs의 출사일수가 3일 정도 빠른 것으로 평가되었다. 따라서 국내에서 배가 반수체 기술을 적용함에 반수체 유기체로서의 TAILs는 사용에 적합할 것으로 판단되었다.
활용 집단의 반수체 유기율 조사
옥수수 집단과 TAILs를 교배하여 생산된 종자의 반수체 유기율은 Table 2와 같다. 2014년에는 미백2호 등 9집단을 이용하였으며 이들 집단의 반수체 유기율은 2.4%-5.3%였다. 2015년에는 JK131 등 13집단을 이용하였으며 이들 집단의 반수체 유기율은 2.5%-6.5%였다. 2016년에는 HW9/HW1 등 16집단을 이용하였으며 이들 집단의 반수체 유기율은 3.4%-9.8%였다. 2017년에는 HW4/15S8025 등 10집단을 이용하였으며 이들 집단의 반수체 유기율은 4.9%-11.8%였다. 2014년부터 2017년까지 총 48집단을 이용하였으며 이들 집단의 반수체 유기율은 2.4%-11.8%로 다양하였다. 연도별 평균을 비교하면 2014년은 3.9%, 2015년은 4.8%, 2016년은 6.7%, 그리고 2017년은 8.3%로 반수체 유기율의 연차간 유의한 차이가 인정되었다. 찰옥수수(미백2호 등 12집단)와 종실용 옥수수(강일옥 등 36집단)로 구분하여 유기율을 비교해본 결과 찰옥수수 유기율은 5.7%, 종실용 옥수수 유기율은 6.9%이었고 통계적 유의성은 인정되지 않았다.
Table 2.
Haploid induction rates in the waxy and field corn populations
| Year |
Population name |
Color expression on seeds (1-5)1) |
Total harvested seeds (g) |
Haploid seeds (g) |
Haploid induction rate (%) |
| 2014 | Mibaek2 | 4 | 12,330 | 353 | 2.9 |
| GW39 | 5 | 3,762 | 92 | 2.4 | |
| W1429 | 4 | 7,687 | 407 | 5.3 | |
| 32P75 | 4 | 6,791 | 271 | 4.0 | |
| P1894 | 5 | 7,126 | 326 | 4.6 | |
| S335 | 5 | 10,199 | 499 | 4.9 | |
| Gwangpyeongok | 4 | 10,512 | 412 | 3.9 | |
| Gangilok | 4 | 9,491 | 391 | 4.1 | |
| MG#4 | 3 | 6,235 | 215 | 3.4 | |
| mean | 4.2 ± 0.72) | 8,237 ± 2622 | 329 ± 122 | 3.9 ± 0.9 | |
| 2015 | NK6240 | 5 | 10,868 | 536 | 4.9 |
| P3502 | 3 | 6,592 | 319 | 4.8 | |
| NK6240/AF05 | 4 | 3,958 | 98 | 2.5 | |
| 32P75/Gangilok | 5 | 7,505 | 490 | 6.5 | |
| P1543/Gwangpyeongok | 4 | 9,339 | 331 | 3.5 | |
| G29/GP | 4 | 18,978 | 511 | 2.7 | |
| G30/Gangilok | 5 | 8,135 | 420 | 5.2 | |
| G31/S335 | 4 | 8,050 | 247 | 3.1 | |
| G32/Gwangpyeongok | 4 | 13,180 | 861 | 6.5 | |
| G32/32P75 | 3 | 9,767 | 575 | 5.9 | |
| JK131 | 5 | 9,988 | 566 | 5.7 | |
| JK147 | 4 | 7,994 | 376 | 4.7 | |
| JK129 | 5 | 8,638 | 547 | 6.3 | |
| mean | 4.2 ± 0.7 | 9,461 ± 3602 | 452 ± 188 | 4.8 ± 1.4 | |
| 2016 | HW9/HW1 | 4 | 6,485 | 221 | 3.4 |
| HW9/14S8015 | 4 | 7,718 | 448 | 5.8 | |
| HW9/HW11 | 5 | 7,145 | 580 | 8.1 | |
| 14S8018/HW3 | 3 | 8,590 | 570 | 6.6 | |
| P1543/Sirus | 4 | 9,467 | 691 | 7.3 | |
| P1543/P154 | 5 | 8,440 | 620 | 7.3 | |
| Gwangpyeongok/P3156 | 4 | 6,520 | 304 | 4.7 | |
| S335/P1184 | 4 | 7,675 | 565 | 7.4 | |
| G35/NK6240 | 5 | 7,080 | 475 | 6.7 | |
| P7709 | 5 | 9,414 | 507 | 5.4 | |
| P8400 | 4 | 7,964 | 449 | 5.6 | |
| 09S5221/07S8040 | 4 | 6,966 | 526 | 7.6 | |
| G30 | 5 | 14,971 | 1,461 | 9.8 | |
| G26 | 3 | 4,796 | 251 | 5.2 | |
| 15S6339/15S6067 | 5 | 16,707 | 1,347 | 8.1 | |
| 15S8058/15S6067 | 4 | 9,339 | 724 | 7.8 | |
| mean | 4.3 ± 0.7 | 8,705 ± 3297 | 609 ± 360 | 6.7 ± 1.4 | |
| 2017 | WP09(FS)A | 4 | 2,928 | 264 | 9.0 |
| WP09(FS)B | 4 | 3,386 | 318 | 9.4 | |
| WPE | 4 | 2,554 | 302 | 11.8 | |
| HW9/16germplasm | 3 | 8,167 | 427 | 5.2 | |
| HW3/16germplasm | 4 | 7,580 | 430 | 5.7 | |
| HW4/15S8025 | 5 | 2,998 | 352 | 11.7 | |
| G29/NK6240 | 5 | 5,731 | 282 | 4.9 | |
| P8521 | 5 | 4,391 | 251 | 5.7 | |
| 32W86/P8521 | 5 | 12,062 | 1,222 | 10.1 | |
| 32P75/P7709 | 5 | 17,676 | 1,599 | 9.0 | |
| mean | 4.4 ± 0.7 | 6,747 ± 4882 | 545 ± 469 | 8.3 ± 2.7 | |
| P value of year3) | - | - | - | < 0.001 | |
반수체 유기율은 집단 특이적으로 작용하고(Prigge et al., 2011), 색소 발현 저해인자의 존재 유무가 반수체 유기율에 영향을 미친다고 하였다(Rober et al., 2005). 본 연구에서 활용한 집단의 유기율 중 가장 낮은 집단은 GW39로 2.4%였고, 가장 높은 집단은 WPE로 11.8%였다. 이는 Prigge 등(2011)과 Rober 등(2005)이 보고한 바와 같은 원인으로 다양한 유기율을 보인 것으로 판단된다. TAILs의 평균 반수체 유기율은 약 6-9%로 보고되었는데(Chaikam et al., 2016) 본 연구에서 4년간 반수체 유기율의 전체 평균은 6.0%로 유사한 유기율을 보였다. 연차 간 반수체 유기율을 비교하면 2014년에는 낮은 유기율은 보였으나, 그 이후 유기율이 증가하는 경향이었다. 배가 반수체 기술을 도입한 초기(2014년-2015년)에 유기율이 낮은 것은 반수체 종자 선별의 미숙함이 어느 정도 작용했을 것으로 판단된다. 또한 온도, 일조 등 연차 간 환경적인 변이도 종자의 색발현에 관여했을 것으로 판단된다.
염색체 배가 시기 비교
반수체 식물체의 염색체 배가에 적합한 생육시기를 평가하기 위하여 염색체 배가 처리시기를 발아종자와 유묘 단계에서 실시하였으며 이에 따른 결과는 Table 3과 같다. P7709 집단에서 발아종자 처리구의 수확계통수는 6, 정식주 대비 수확계통수의 비율을 나타내는 계통생산율은 3.8%였고, 유묘 처리구의 수확계통수는 42, 계통생산율은 13.1%였다. P8400 집단에서 발아종자 처리구의 수확계통수는 9, 계통생산율은 4.5%였고, 유묘 처리구의 수확계통수는 42, 계통생산율은 12.4%였다. 그리고 14S8018/HW3 집단에서 발아종자 처리구의 수확계통수는 45, 계통생산율은 5.4%였고, 유묘 처리구의 수확계통수는 32, 계통생산율은 8.4%였다. 옥수수 3집단에 대하여 발아종자와 유묘 처리구의 평균을 비교하면 발아종자 처리의 계통생산율은 4.5%, 유묘 처리의 계통생산율은 11.3%로 처리별 유의한 차이가 인정되었다.
Table 3.
Comparison of the chromosome doubling stage in three different populations
| Doubling stage1) | Population |
No. of transplants (plants) |
No. of self pollinated (plants) |
No. of harvested ears (lines) |
Production rate (%) |
| Germinated seeds | P7709 | 160 | 35 | 6 | 3.8 |
| P8400 | 200 | 43 | 9 | 4.5 | |
| 14S8018/HW3 | 840 | 100 | 45 | 5.4 | |
| mean | 400 | 59 | 20 | 4.5 | |
| Seedlings | P7709 | 320 | 116 | 42 | 13.1 |
| P8400 | 340 | 136 | 42 | 12.4 | |
| 14S8018/HW3 | 380 | 89 | 32 | 8.4 | |
| mean | 347 | 114 | 39 | 11.3* |
배가 반수체 기술을 옥수수 육종에 적용함에 있어 염색체 배가는 화분 발생 및 수정과 밀접한 연관이 있으므로 염색체 배가 효율을 높이는 것이 인공교배의 성공 효율을 높일 수 있는 방법이다. 염색체 배가 처리시기를 발아종자와 유묘 단계에서 비교한 결과 유묘 단계에서 처리하는 것의 효율성이 높은 것으로 평가되었다. 발아종자 단계는 매우 연약한 단계이지만 colchicine 용액에 12시간 침지해야 하고, 작업 과정에서의 상처 및 절단 등의 피해가 발생할 가능성이 높은 것이 낮은 계통생산율에 관여했을 것으로 판단된다.
배가 반수체 기술에 의해 육성된 계통의 농업형질 평가
배가 반수체 기술을 이용하여 육성한 계통의 농업형질을 평가한 결과는 Table 4와 같다. 종실용 옥수수 육성 5계통(17DHL01-17DHL05)의 출사일수는 77-79일로 대비계통 HF1과 유사하였고, 도복은 1.7 이하로 내도복성이었다. 간장 및 착수고는 대비계통과 유사하거나 큰 경향이었고, 이삭장은 10.4-15.5 cm로 다양하게 분포하였다. 17DHL01-17DHL03의 100립중은 대비계통보다 4-5 g 무거운 특성을 보였다. 찰옥수수 육성 5계통(17DHL07-17DHL11)의 출사일수는 71-75일로 대비계통 HW9보다 빨랐고, 도복은 1.3 이하로 내도복성이었다. 간장은 대비계통보다 큰 경향이었고, 이삭 폭은 대비계통과 유사하거나 좁은 경향이었다.
Table 4.
Agronomic characteristics of selected doubled haploid (DH) lines at Hongcheon in 2017
| DH lines |
Days to silking (days) |
Lodging (1-9) |
Plant height (cm) |
Ear height (cm) |
Ear length (cm) |
Ear width (cm) |
Ear row (rows) |
100 seeds weight (g) |
Kernel types |
| 17DHL011) | 78 | 1.0 | 205 | 55 | 13.1 | 3.8 | 11.3 | 36.7 | Semi-dent |
| 17DHL02 | 78 | 1.0 | 202 | 64 | 11.5 | 3.5 | 12.0 | 35.4 | Dent |
| 17DHL03 | 77 | 1.3 | 212 | 54 | 13.5 | 3.5 | 12.0 | 36.2 | Intermediate |
| 17DHL04 | 77 | 1.3 | 214 | 82 | 15.5 | 3.7 | 13.3 | 26.5 | Flint |
| 17DHL05 | 79 | 1.7 | 189 | 51 | 10.4 | 4.2 | 14.0 | 31.9 | Semi-dent |
| HF1 | 77 | 2.0 | 193 | 51 | 12.9 | 3.6 | 12.0 | 31.4 | Intermediate |
| P value | < 0.05 | ns | ns | ns | ns | < 0.05 | ns | < 0.001 | |
| 17DHL072) | 71 | 1.3 | 178 | 65 | 10.9 | 3.0 | 11.3 | 28.8 | Waxy corn |
| 17DHL08 | 75 | 1.0 | 153 | 56 | 11.0 | 3.0 | 11.3 | 26.2 | Waxy corn |
| 17DHL09 | 75 | 1.0 | 142 | 50 | 13.6 | 3.0 | 10.7 | 23.0 | Waxy corn |
| 17DHL10 | 74 | 1.0 | 158 | 53 | 12.1 | 3.1 | 12.0 | 22.2 | Waxy corn |
| 17DHL11 | 74 | 1.3 | 136 | 45 | 12.0 | 3.3 | 12.0 | 24.6 | Waxy corn |
| HW9 | 79 | 1.0 | 129 | 47 | 12.0 | 3.3 | 12.0 | 24.7 | Waxy corn |
| P value | < 0.001 | ns | ns | ns | ns | ns | ns | < 0.01 |
본 연구에 활용한 육성 10계통은 예비 선발을 거친 계통이므로 옥수수 육종에 활용하기에 모두 적합한 계통으로 평가되었다. 특히 종실용 옥수수에서 17DHL01, 찰옥수수에서 17DHL10 등은 식물체의 초형 및 이삭 형태 등을 고려할 때 농업형질이 매우 우수한 것으로 판단된다. 따라서 이 선발 계통을 교배조합의 모, 부본으로 사용하여 교잡종을 만들고 이를 평가하면 우량한 교잡종의 선발이 가능할 것으로 판단된다.
결 론
옥수수 품종육성은 계통육성과 육성된 계통을 이용한 교잡종의 선발단계로 구분할 수 있다. 계통육성은 우량한 교잡종을 만들기 위해 필수적으로 수행해야 하는 과정이며 국내에서 계통육성은 전적으로 전통적인 육종방법에 의존하고 있다. 이러한 전통육종방법은 지루하고 시간이 많이 소요되는 과정이다. 옥수수에서 배가 반수체 기술은 단기간에 계통을 육성할 수 있는 기술이며 해외 선진 옥수수 연구기관 및 회사에서는 이 기술을 적극적으로 활용성하고 있다. 본 연구는 배가 반수체 기술을 도입하고 옥수수 육종시스템으로 정착시키고자 수행하였다. 옥수수와 교배하여 반수체를 유기할 수 있는 반수체 유기체(inducer)는 배가 반수체 기술의 도입에 필수적인 사항이며, 옥수수연구소는 반수체 유기체인 TAILs의 상업적 사용권을 확보하였다. 국내에서 아열대형 유기체인 TAILs의 출사일수는 72일이었으며 개화기가 국내 옥수수 집단과 교배하기에 적당하였다. 48 집단과 유기체의 교배를 통하여 생산된 종자에서 반수체 유기율을 평가한 결과 평균 6.0%였다. 2014년부터 2017년까지의 반수체 유기율은 각각 3.9, 4.8, 6.7, 8.3%로 나타났다. 반수체 종자의 염색체 배가 처리시기를 비교해본 결과 유묘 단계에 처리하는 것이 발아묘 단계에 처리하는 것보다 효율성이 우수하였다. 배가반수체 기술에 의해 육성된 옥수수 계통의 농업 형질을 평가하였으며 옥수수 육종에 활용하기에 모두 적합한 계통으로 판단되었다. 반수체 유기율의 향상 및 반수체 종자 선별효율 증대 등 개선이 필요한 부분도 있지만, 배가 반수체 기술의 활용으로 국내의 옥수수 육종 효율 및 능력은 크게 향상될 것으로 기대된다.
