

서 론
재료 및 방법
연구대상지
조사분석
결과 및 고찰
수리・수문학적 특성
계절에 따른 수질변화 특성
동물플랑크톤 군집구조의 시・공간적인 변동
Daphnia 출현과 투명도와의 상관관계
요 약
서 론
도암호 유역의 고랭지 경작은 목축업이 위축되기 시작한 1970년대 이후 고랭지농업이 지역의 기반산업으로 자리매김하면서 활발하게 이루어져 왔다. 고랭지농업의 경우 매년 반복적인 객토와 다량의 비료 투입이 필요한 경작 특성을 갖고 있을 뿐만 아니라 작물의 재배 기간이 짧고 나대지 상태로 방치되어 있는 기간이 긴 특성을 보인다.
우리나라는 몬순기후지역에 속하기 때문에 강우의 약 70%가 7월에서 9월 사이에 집중된다. 이 시기는 작물을 수확하고 난 후 경작지가 나대지 상태로 방치되어 있는 시기와 일치하기 때문에 강우 시 인근 하천으로 다량의 토사가 유입되는 문제점이 있다.
하천과 같은 유수생태계에서 탁수 발생으로 인해 수체가 혼탁해지는 기간은 물의 지속적인 흐름으로 인해 짧아 수서생물에게 미치는 영향은 크지 않은 반면, 하천 바닥으로 침강되어 퇴적되는 토사의 경우에는 특히, 저서성 생물서식지를 훼손하여 저서성 생물에게 부정적이고 만성적인 문제로 작용할 수 있다. 한편, 지리적으로 유수생태계와 유수생태계 사이에 건설된 인공호의 경우, 호내로 유입하는 하천수와 호내 수온의 수직분포 중 일치하는 수온층으로 관입 이동하여 댐 방류구를 통해 하류하천으로 방류되는 수리수문학적인 특성이 있다. 이들 중층방류를 하는 인공호에서 강우 시 관입한 탁수층은 호내 일차생산자의 증식 및 종변화를 촉진시키는 영양염류의 공급원이 되고, 상위소비자인 동물플랑크톤, 어류로 이어지는 에너지 흐름을 좌우하는 역할을 한다(Kim et al., 2018; Lee et al., 2013a; Lee et al., 2013b). 특히, 탁수의 유입은 호소의 부영양화를 촉진시키는 요인으로 작용하여 유해성남조류의 증식 또는 상위소비자로 이어지는 에너지 흐름을 변화시켜 동물플랑크톤 군집구조에 영향을 미칠 수 있다(Chen and Chen, 2017; Shin et al., 2008).
국내 대부분의 호소 또는 인공댐은 특히, 몬순기의 강우 강도를 고려하여 인위적으로 수문개폐를 조절하기 때문에, 호내 탁수층의 형성 시 이동경로 및 체류시간 등은 동물플랑크톤 군집구조 특성 및 변화를 이해하는데 중요하다(Kim et al, 2018; Lee et al., 2013b).
도암호는 2001년 이후 운영이 중단된 상황에서 2022년 현재까지 표층수만이 자연 월류되고 있다. 도암호의 수리수문학적인 특성 때문에 관입된 탁수층은 즉시 방류되지 못하고 호내에 장기간 체류하거나 수온성층이 소멸되는 시기에는 전 수층으로 탁수가 확산되어 동물플랑크톤 군집에 지속적인 영향을 미치는 것으로 판단된다. 이와 같이 연중 탁수에 의한 영향에 노출되어 있는 도암호는 탁수로 인한 수환경변화 및 동물플랑크톤 군집을 파악하기 위한 연구지로써 최적의 환경이라 판단된다.
동물플랑크톤은 일차생산자와 상위소비자의 에너지를 전달하는 매개자일 뿐만 아니라 수생태계의 환경변화에 대한 지표생물로써의 역할을 담당하는 중요한 분류군이다(Ji et al., 2013). 호소생태계의 영양단계에 따라 동물플랑크톤 군집은 일차생산자인 식물플랑크톤 또는 microbial loop와 밀접하게 연계되어 있을 뿐만 아니라 이차소비자인 어류의 중요한 먹이원으로써 어류의 생산성을 좌우하는 중요한 역할을 담당한다(Lee et al., 2010; Noh and Han, 2008). 일반적으로 몬순지역의 온대호소에서 동물플랑크톤 군집은 계절변동에 따라 현존량 증감을 보이고, 이와 같은 양상은 인공호의 수질환경 이외에도 생물상호작용 그리고 수리수문학적인 요인 등과 관련성이 있는 것으로 알려져 있다(You et al., 2010). 동물플랑크톤 군집에 대한 구조적인 특성은 정수생태계의 영양상태(trophic status)를 반영하여 수질특성 변화에 대한 지표로써 사용되기도 한다(Sangakkara and Wijeyaratne, 2015; Sládeček, 1983; Odds et al., 2014).
국내・외에서 동물플랑크톤 군집과 관련한 연구는 주로 호소의 부영양화와 관련하여 다루어져 왔고, 동물플랑크톤 군집의 서식환경에 따른 종조성의 차이를 파악하고자 하는 연구도 이루어진 바가 있다(Choi et al., 2012; Ismail and Adnan, 2016; Lee et al., 2010). 또한 국내 인공호를 대상으로 한 동물플랑크톤 군집과 관련한 연구는 시・공간적인 분포와 변동을 수질환경의 변화, 몬순강우 시 방류패턴으로 인한 영향, 어류의 포식압, 계절에 따른 식물플랑크톤의 종조성의 변화로 인한 먹이원의 질적 양적인 특성 등의 원인과 결부시킨 연구가 있다(Kim et al., 2018; Lee et al., 2013a; Lee et al., 2013b).
탁수가 호소생태계에 미치는 영향에 대한 연구는 몬순강우 시에 국한되어 있거나 유역에서의 탁수유입과 호소의 부영양화 간의 관계를 파악하고 수질환경 개선을 위한 방안을 수립하는데 활용되어 왔다(Nunes et al., 2022).
그러나 송천유역을 대상으로 수질변화에 대한 지속적인 모니터링이 이루어지고 있음에도 불구하고 실질적인 개선 유무에 대한 평가는 불명확한 상황이 지속되고 있다. 수질 모니터링을 통한 수생태계의 환경해석은 매년 유역의 토지이용변화 및 강우패턴에 의존적이고, 연중 일부 특정 시기에 국한된 수질정보 위주의 자료만을 제공하는 반면, 수질환경변화뿐만 아니라 생물서식환경에 대한 장단기적으로 누적된 정보를 제공해 주는 수서생물을 이용한 연구결과는 부족한 상황이다. 최근까지도 국내에서는 연중 또는 지속적으로 호내에 체류하는 탁수의 영향을 받는 호소생태계에서 탁수가 수서생물 군집에 미치는 영향 특히, 동물플랑크톤 군집구조에 미치는 영향 및 변화특성을 규명한 사례는 드물다.
따라서, 본 논문은 도암호 상류 송천유역에 분포하는 고랭지경작지에서 유역하는 탁수의 영향에 빈번하게 노출되어 있는 호소생태계에 대한 수서생물 군집의 반응변화를 이해하고 관리 대책을 마련하기 위한 기초연구의 일환으로, 현재 표층 월류방식에 의존하고 있는 도암호를 대상으로 동물플랑크톤 군집의 구조적인 특성변화를 파악하고자 2016년~2018년까지 12회에 걸쳐 동물플랑크톤의 출현종 및 현존량을 조사・분석하였다.
재료 및 방법
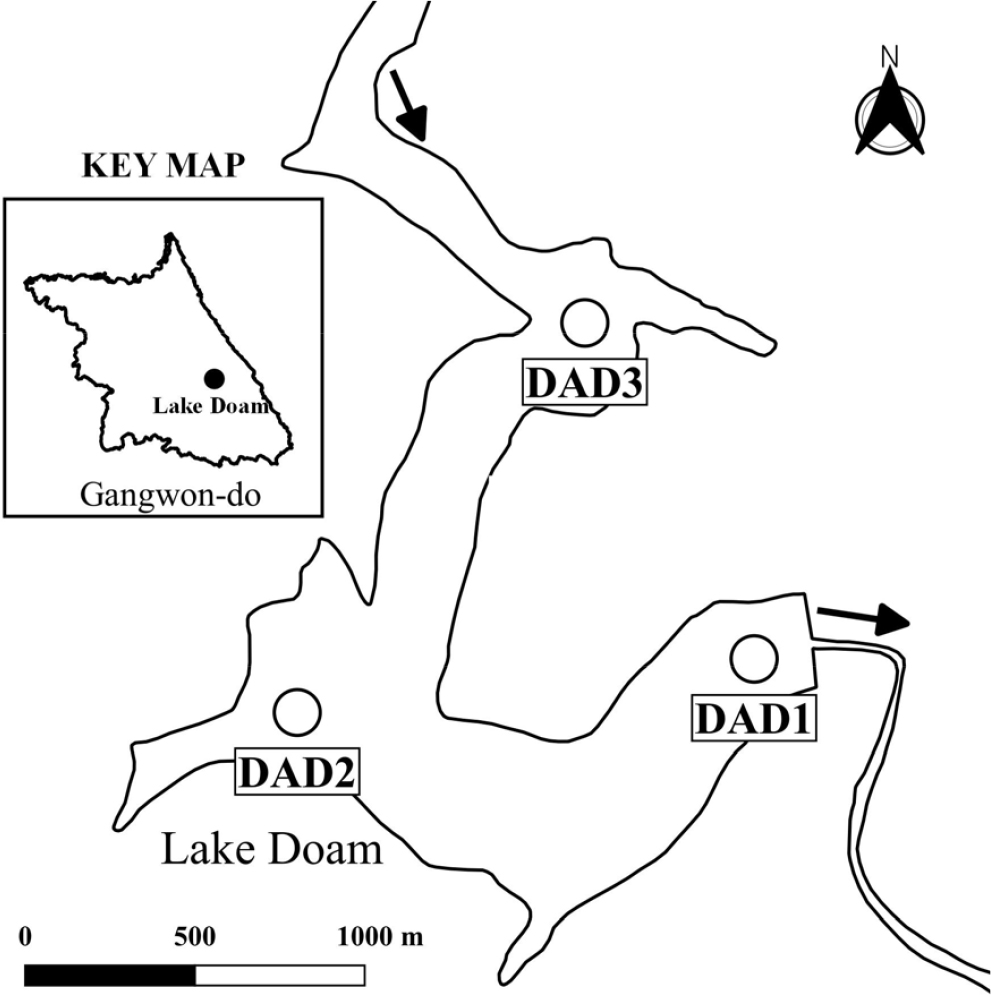
연구대상지
도암호는 강원도 평창군 도암면 수하리에 위치하고 있으며, 유역변경식 수력 발전을 위해 남한강 상류인 송천을 막아 식수원공급을 목적으로 1991년 건설된 인공호로 유역면적은 144.9 km2이며, 평균저수량이 51 × 106 m3, 유효저수량이 4 × 107 m3이다(Fig. 1). 송천의 상류 고랭지농업지역에서 발생하는 탁수의 영향으로 수질이 악화된 이후부터 현재까지 운영이 중단된 상태로 표층을 통해 월류식 방류를 하고 있다. 도암호 유역은 고랭지 밭에서의 토사 유출과 비점오염발생으로 인한 수질악화의 영향으로 2007년에 환경부에서는 비점오염원 관리지역으로 지정하고 있다.
조사분석
조사는 매년 유출량이 안정적이고 낮은 시기에 수행하였고 유입하천의 영향을 가장 크게 받는 유입 지점(DAD3), 호심 지점(DAD2) 그리고 댐앞 지점(DAD1) 등 3개소에서 수행하였다. 동물플랑크톤을 채집하기 위해 선상에서 62 µm mesh의 동물플랑크톤 넷트를 수직으로 5 m 수심까지 내린 후 1 m/sec로 끌어 올렸다. 넷트로 농축한 시료는 100 ml 투명 bottle에 담아 최종농도가 4% formalin이 되도록 고정한 후 실험실로 운반하였다. 검경을 위해 농축시료에서 1ml의 subsample을 얻어 Sedgewick-Rafter cell chamber로 옮긴 후 광학현미경(Nikon Eclipse Ci) 하에서 출현 종을 분류군별로 계수하였고 계수된 동물플랑크톤은 ind./L로 환산하여 현존량으로 제시하였다.
동물플랑크톤 군집의 계절변동과 더불어 현장 및 실험실에서 각각 수질항목을 분석하였다. 수질과 관련한 조사항목은 수온, 투명도, 탁도, 부유물질(SS), 총인(TP), 총질소(TN) Chl-a 등으로, 현장조사 항목 외에는 분석을 위해 채취한 시료를 암상태로 냉장운반하여 Ministry of Environment(2017)에 근거하여 분석하였다.
결과 및 고찰
수리・수문학적 특성
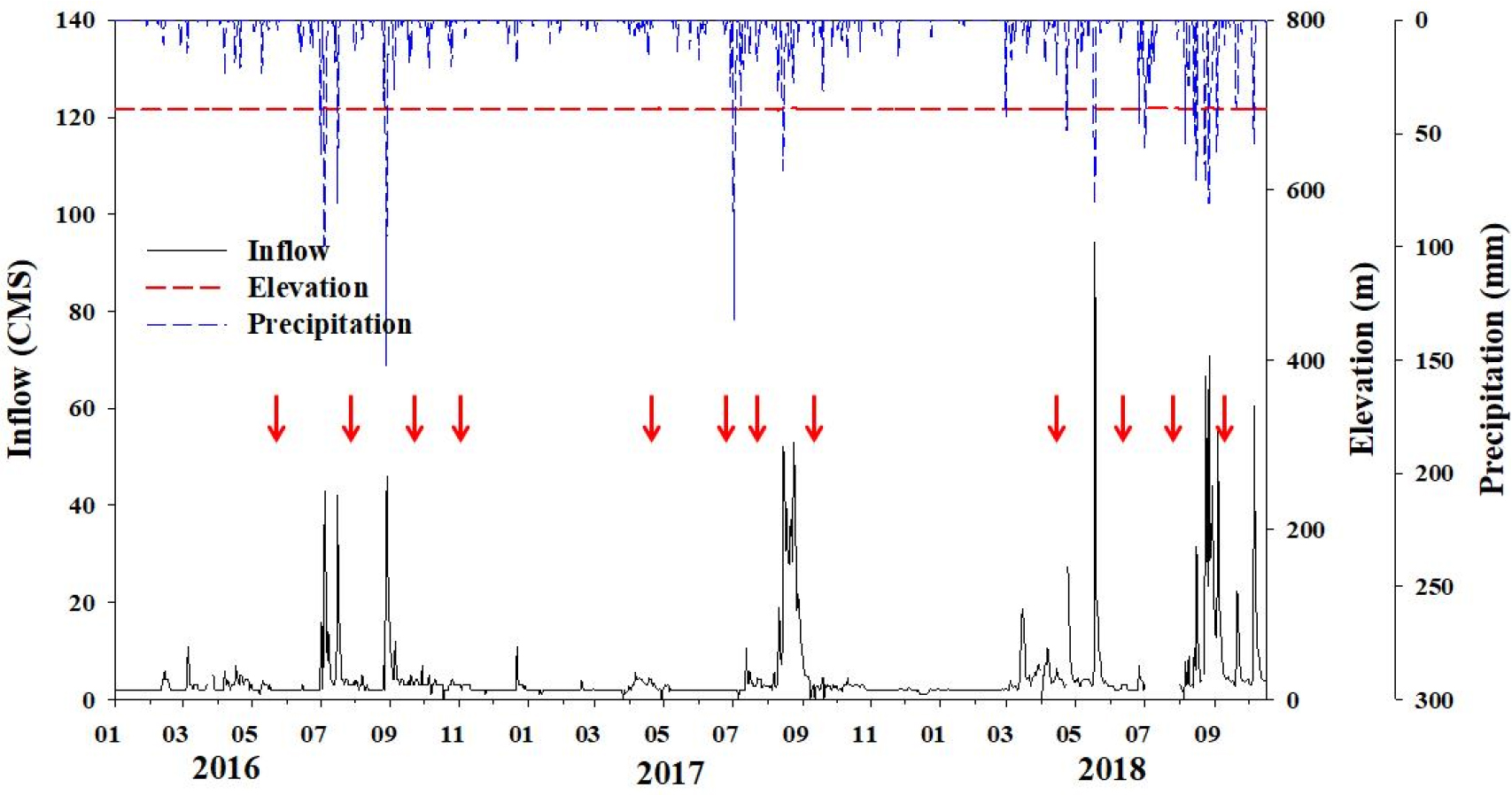
도암호 유역의 총 강수량은 조사 기간 2017년에 1,025 mm로 가장 적었고, 2018년에는 1,730 mm으로 가장 많았다(Fig. 2). 또한 2018년에는 강우의 빈도가 이전 연도들과 달리 산발적이고 지속적으로 내렸다. 시기별 강수량의 변동은 7-9월에 전체 강우량의 66.6%가 집중되었다. 조사 기간 유입수량을 비교한 결과, 2016년이 가장 적은 유입수량을 보였고 매년 7월과 9월에 반복적인 강우로 인해 호내로의 유입수량이 공통적으로 급증하는 양상을 보인 반면, 2018년에는 초여름에도 호내로 유입하는 수량이 많았다. 7-9월의 총 유입수량과 총 유출수량은 각각 53%, 52%를 차지하였다. 도암호 평균 수위는 694(± 11) m이었으며, 연간 수위의 변동은 크지 않았다.
계절에 따른 수질변화 특성
조사기간, 강우시기에 송천을 통해 유입되는 탁수는 호내 부유물질 농도를 높이고 동시에 총인의 농도 역시 높이는 결과를 보였다(Table 1). 이와 같은 결과는 도암호 상류 유역 하천들과 호내의 수질특성을 분석한 결과와 일치한다(Kwon et al., 2020; You et al., 2010).
Table 1.
Mean and standard error values of the physicochemical characteristics of water in Lake Doam from April 2016 to December 2018 (WT, water temperature; DO, dissolved oxygen; Turb, turbidity; Chl-a, chlorophyll a; SS, suspended solid; TP, total phosphorus; TN, total nitrogen)
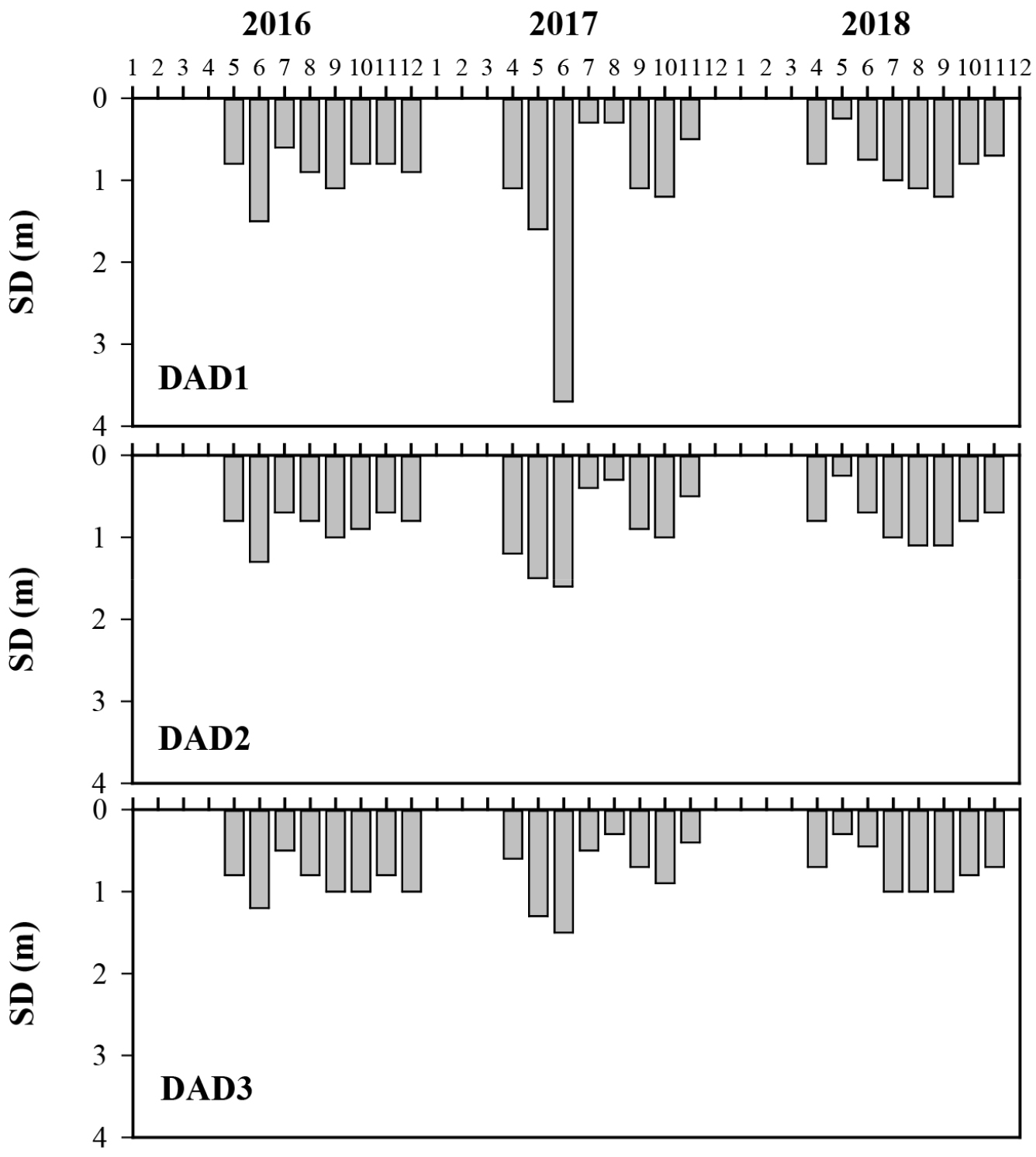
도암호는 일순환호로, 5월에서부터 10월 사이에 수온성층이 유지되고 11월 이후 소멸하여 수체 전체가 혼합된다(Fig. 3). 도암호에서 수온은 2016년 4.1°C-26.6°C(10.0 ± 6.0°C), 2017년 1.5°C-22.1°C(8.4 ± 5.6°C), 2018년 0.0°C-27.3°C(10.4 ± 6.3°C)의 범위에서 수직 및 수평변동을 보였다. 2017년과 2018년의 경우 특히, 2018년에는 유입하천에서 댐앞 지점(DAD1)의 바닥층까지 관입된 높은 수온층이 확인되었다. 투명도는 0.3-3.7 m(0.9 ± 0.5)의 범위로, 지점별 변동 패턴에는 차이가 없고, 유입 지점(DAD3)에 비해 댐앞 지점(DAD1)에서 다소 높아지는 양상을 보였다(Fig. 4). 이는 도암호에 유입된 탁수입자는 대부분 유입부에서 침강되기 때문으로 판단된다. 결과적으로, 조사 기간 도암호에서 높은 투명도를 보이는 시기는 유입수량이 적고 수위의 변동이 크지 않아 수체가 안정화되어 있는 시기로 판단된다. 특히 2017년 6월에는 댐앞 지점(DAD1)에서 3.7 m로 가장 높은 투명도를 보였다(Fig. 4).

부유물질 농도는 0.1-631.0 mg/L의 범위이었으며 평균 28.2(± 58.9) mg/L이었다(Fig. 3). 100 mg/L 이상의 높은 부유물질 농도는 조사 기간 5 m 이심에 주로 형성되었다. 도암호 내 높아진 부유물질농도는 댐월류에 의한 소멸보다는 탁수의 유입이 감소하고 수온성층이 소멸되는 10월 이후에 수체순환 및 유입수로 인한 희석에 의해 농도가 낮아지는 양상을 보였다.
부유물질농도의 지점별 수직 및 수평변동을 비교해 보면, 유입 지점(DAD3)에서는 높은 부유물질농도가 시기별 강우패턴에 따라 표층 또는 호소 바닥으로 확산하였다. 부유물질농도는 유입 지점(DAD3)에서 댐앞 지점(DAD1)까지 이동하는 동안에 전 수층으로 확산하였는데, 높은 부유물질농도의 경우, 표층에 비해 중층 이심에서 주로 확인되었다. 이는 표층 월류방식의 도암호에서는 유입되는 굵은 입자의 부유물질이 호내에 지속적으로 퇴적되고 있음을 시사한다.
탁수층의 호내 체류 지속기간은 바닥층을 제외하고 호내 5 m 이심으로 유입 후 2016년과 2017년에는 7월-8월 사이 2개월 동안 호내에 체류하며 전수층으로 확산하였다. 반면, 2018년에는 5월에서 9월까지 5개월에 걸쳐 전 수층이 고탁수에 노출되어 있었다. 2018년 5월 수층별 부유물질농도는 55.2 mg/L(표층)-490.2 mg/L(30 m)의 높은 농도를 보인 후 9월까지 고탁수가 유지되었고, 10월에 이르러서야 12.5 mg/L(표층)-20.2 mg/L(30 m) 범위까지 감소한 것으로 확인되었다.
본 연구에서 부유물질 농도는 총인농도 및 Chl-a농도를 증가시키는 요인이라 판단된다(Fig. 3). TP(총인)농도는 0.010-0.587 mg/L의 범위로 평균 0.075(± 0.088) mg/L이었고, Chl-a농도는 0.04-201.12(11.64 ± 17.91 mg/L)의 범위를 보였다(Fig. 3). Chl-a농도는 강우로 인한 탁수 유입에 의한 부유물질 농도가 증가 이후 0 m-5 m 사이에서 증가하는 양상을 보였다. 한편 Chl-a농도의 수직적인 분포는 하천과 인접해 있는 유입 지점(DAD3)과 호심 지점(DAD2)의 경우에는 수직적으로 높은 Chl-a농도가 patch를 형성하였다. 이는 하천의 상류에서 탈리되어 호내로 유입된 부착조류의 기여가 있는 것으로 판단된다.
결과적으로, 송천을 통해 도암호로 유입되는 탁수는 호내 영양염류(특히 총인)의 공급원이 되고 있고 수직적으로 높은 Chl-a농도의 patch를 형성하는 원인으로 보인다(Kwon et al., 2020; Nunes et al., 2022; You et al., 2010). 이와 같은 도암호의 높은 표층 Chl-a농도를 보이는 시기에는 동물플랑크톤 군집의 구조적인 변화에 영향을 주는 중요한 요인이라 판단된다.
동물플랑크톤 군집구조의 시・공간적인 변동
전 조사 기간 및 지점에서 도암호의 동물플랑크톤 군집은 총 48종 5,780.5 ind./L의 출현이 확인되었으며, 분류군별 출현종 수 및 현존량의 상대적인 점유율은 각각 지각류가 8.3%, 6.7%, 요각류 16.7%, 13.1%, 원생동물 6.3% 11.1% 그리고 윤충류가 68.8%, 69.2%로, 도암호의 동물플랑크톤 군집구조는 윤충류의 점유율이 높은 호소생태계로 파악되었다(Fig. 5). 조사기간 누적된 연도별 총 출현종 수와 현존량은 2016년에 29종(8종-19종) 1,942.3 ind./L(70.9 ind./L-374.1 ind./L), 2017년에 34종(2종-18종) 2,852.6 ind./L(47.2 ind./L-906.5ind./L) 그리고 2018년에는 25종(5종-15종)종 985.5 ind./L(10.4 ind./L-312.5 ind./L)으로, 조사 기간 중 높은 강우빈도의 영향을 받은 2018년에 가장 적은 출현종 수 및 현존량을 보였다(Fig. 5).
기존의 연구들에서는 일반적으로, 하천을 막아 형성된 인공호는 몬순강우 시기에는 수체의 교란이 빈번해지기 때문에 동물플랑크톤 군집구조는 대형동물플랑크톤보다 소형동물플랑크톤인 윤충류에 의해 우점하는 군집구조를 이루는 것으로 알려져 있다(Beaver et al., 2013). 또한 부영양화된 호소생태계에서도 소형동물플랑크톤인 윤충류에 속하는 종들이 군집구조에서 우점하는 종으로 확인되고 있다(Kim et al., 2000; Kim et al., 2005).
본 연구에서는 도암호 역시 부영양호에 속하는 호소생태계로, 동물플랑크톤 군집구조는 연중 도암호내에 체류하는 탁수입자의 영향을 받아 작은 크기 위주의 종 조성을 이루는 양상을 보였는데, 도암호 상류 유역의 토지이용에 따른 영향과의 관련성에 대해서는 추가적인 검토가 필요하다.
도암호에서 동물플랑크톤 군집의 출현종 수와 현존량은 시・공간적으로 매년 비슷한 변동 양상을 보였는데, 6월 또는 7월에 비교적 높았고, 수온이 낮아지는 춘계(4월) 또는 추계(9월)에는 낮은 경향을 보였다(Fig. 5). 특히, 현존량의 경우, 강우 직후 현저한 감소패턴이 뚜렷하였다(Fig. 5). 이와 같은 결과는 몬순지역의 일순환 호소생태계에서 일반적으로 나타나는 동물플랑크톤 군집의 계절변동과 일치하는 결과로, 이는 물리적인 씻김현상 때문으로 알려져 있다. 2017년 6월에는 동물플랑크톤의 현존량이 높은 시기인 반면에, 2017년 7월 강우 직후에는 동물플랑크톤의 현존량이 낮아졌다. 이와 같은 양상은 도암호에서 매년 강우를 전후로 하여 반복적으로 나타나는 현상이고 특히, 2018년에는 빈번한 강우와 유입수량의 증가로 인한 고탁수의 충층유입이 다른 해에 비해 강했던 해이다.
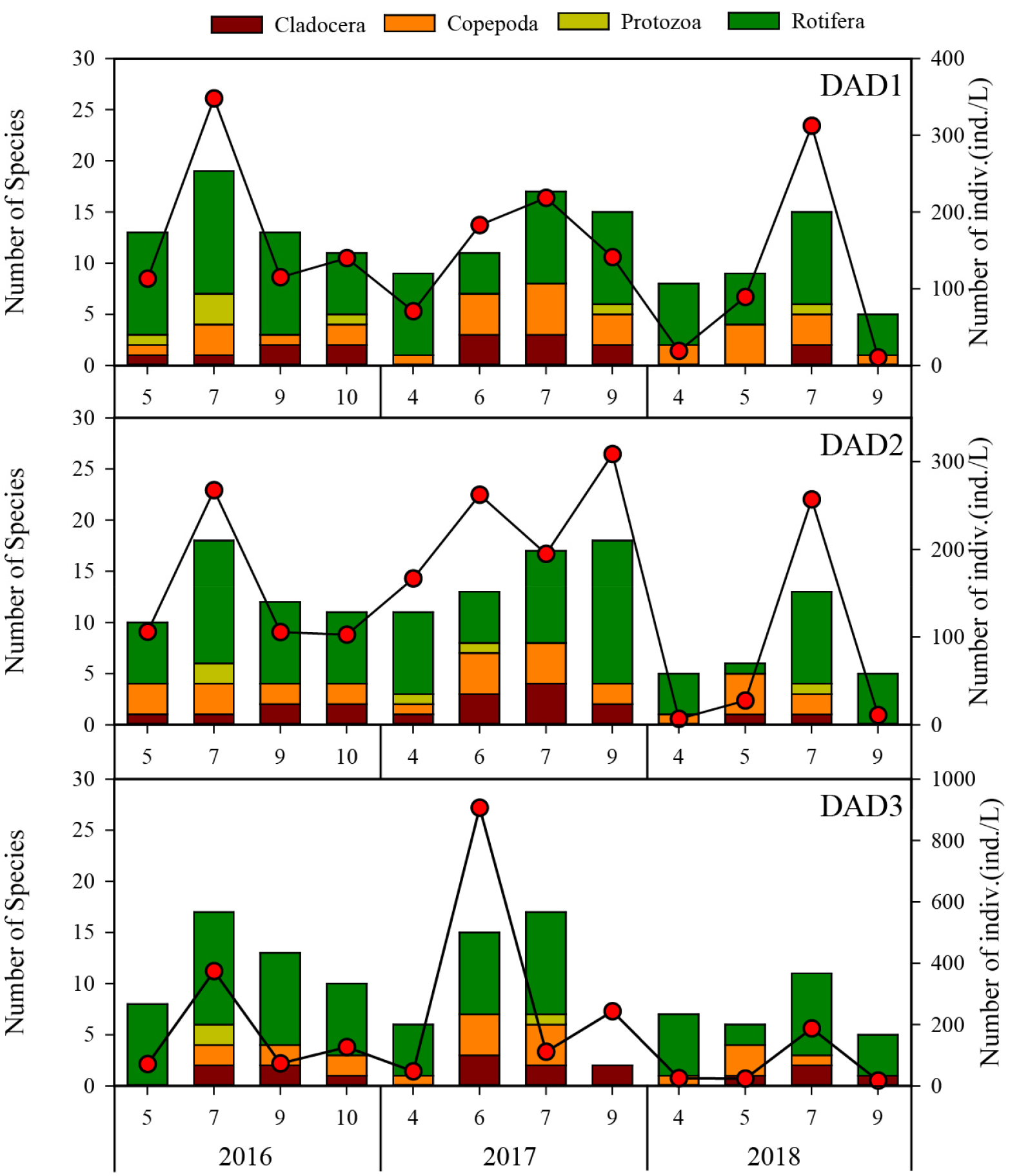
조사 시기 중 지각류에 대한 출현종 수의 점유율은 2017년에 가장 높았으나, 현존량의 점유율은 2016년에 가장 높았고, 동물플랑크톤의 현존량이 높은 시기는 6월 또는 7월에 확인이 되었다. 이 시기에는 도암호에서 수온이 높아지는 시기(19.5°C-27.3°C)이며 투명도가 증가하는 시기이기도 하다. 특히, 2017년 6월에는 빙어가 선호하는 먹이원인 대형지각류(Daphnia galeata)의 현존량 또한 현저하게 높은 시기와 일치하였다.
호소생태계에서 특정 시기에 동물플랑크톤의 현존량이 뚜렷한 증감을 보이는 이유 중 하나는 어류에 의한 포식압이 있다(Li and Zhang, 2015; Oh et al., 2019). 도암호에서 2018년 확인된 어류는 총 14종으로, 비록 계절변화의 영향을 받아 우점종의 조성이 바뀔 수는 있으나, 빙어(53.5%)가 우점하였고, 이 외에 상대풍부도가 10% 이상인 어종으로 참마자, 피라미, 참붕어 등의 부유성 어종이 우세한 호소생태계로 확인되었다.
도암호에서 우점하는 빙어는 동물플랑크톤 특히, 지각류를 주요 먹이원으로 하는 냉수성 어종이다(Chang et al., 2005; Yoshioka, 1991). 도암호에서 서식하는 냉수성 어종인 빙어의 경우, 여름에는 수온이 높은 표층을 피하여 수온이 낮은 수층에 무리를 이루어 분포하기 때문으로 어류의 먹이원으로 적합한 중대형동물플랑크톤을 포식할 수 있는 기회가 많지 않을 수 있다(Oh et al., 2019; Yoshioka, 1991; Yoshioka and Wada, 1994).
잡식성 어종 중 빙어 이외의 부유성 어류의 경우, 빙어와 다른 유광층과 호안에 서식 분포하면서 성장단계에 따라 먹이전환을 하기 때문에 동물플랑크톤 군집에 대해 시・공간적으로 영향을 줄 수 있다(Li and Zhang, 2015). 도암호의 어류군집에서 확인된 부유성 어종은 광온성 어류이고, 대부분의 어종이 시야에 의존하는 먹이 포식을 한다(Rowe et al., 2003).
도암호는 어류의 먹이원인 동물플랑크톤 군집을 구성하는 종의 크기가 주로 100 µm 전후의 소형동물플랑크톤이고, 탁수의 영향에 지속적으로 노출되어 있는 호소생태계이기 때문에, 시야에 의존하여 대형동물플랑크톤(지각류와 요각류)을 주로 먹이원으로 섭이하는 성어에게는 부적합한 먹이환경이라 판단된다.
자연 월류식 방류를 하는 도암호의 경우, 기후환경 변화 및 유역의 수변환경 변화로 인해 수온이 증가하고 고탁수가 중층 이심으로 유입이 이루어지는 상황이 지속된다면 여름동안 낮은 수온층에 무리를 짓는 냉수성 어종인 빙어 개체군은 점진적으로 개체수 및 생체량 감소(왜소화)를 겪을 것으로 예상된다.
호소생태계에서 동물플랑크톤 현존량 증감의 또 다른 원인은, 남조류 증식과의 관련성이다(Sampaio et al., 2002). 도암호에서 Chl-a농도는 7월에 낮고 9월에 높아지는데, 이 기간 식물플랑크톤 군집의 종조성은 녹조류에서 남조류로의 천이를 보인다. 특히, 2018년 9월에는 남조류의 출현 시기가 9월에서 7월로 앞당겨진 해로 확인되었을 뿐만 아니라 유해성남조류(Anabaena spp., Aphanizomenon flos-aquae, Microcystis sp.)가 출현한 시기와 동물플랑크톤의 현존량이 급감소하는 시기와 일치하였다(Wonju Regional Environmental Office, 2018). 몇몇 연구에서 호소생태계에서 부영양화 상태는 동물플랑크톤의 출현종 수 및 현존량을 감소시키는 요인으로 지목된 바가 있다(Kim et al., 2000). 이는 부영양호에서 과다번성한 남조류의 독성이 동물플랑크톤에게 부정적인 영향을 주고 특히, 먹이원인 식물플랑크톤을 여과섭식하는 동물플랑크톤에게 섭식 저해를 야기시키는 원인이 되고 있기 때문이다(Lee et al., 2010; Lee et al., 2010; Lee et al., 2013b; Sampaio et al., 2002).
도암호에 출현하는 Asplanchna priodonta는 남조류를 먹이원으로 하는 동물플랑크톤 종으로 알려져 있다(Pociecha and Wilk-Woźniak, 2008). Asplanchna priodonta가 출현하는 시기에 함께 높은 현존량을 보이는 공서종으로는 Brachionus angularis와 Keratella cochlearis로 확인되었다. Asplanchna priodonta의 현존량은 2018년 6월과 7월 사이에 높은 현존량을 보였고, 9월에는 다른 동물플랑크톤 종들과 함께 급감소하였다. 이는 남조류가 번성하는 9월에 다른 동물플랑크톤 종들의 현존량 감소와 함께 Asplanchna priodonta의 현존량이 급감소 하는 원인이, 단순히 남조류 증식으로 인한 영향을 받았기 보다는 9월 초 유입수량의 급증으로 인한 물리적인 씻김효과가 오히려 큰 것으로 사료된다.
국내 인공호에서 수체의 체류시간은 강우패턴에 의존하고 있고 인공호에서의 방류조절은 동물플랑크톤의 현존량 변화를 좌우하는 중요한 수문학적인 요인으로 알려져 있다. 동물플랑크톤 군집에서 현존량의 증감이 수위조절과 밀접한 관련성이 있다는 보고는 일반적이다(Beaver et al., 2013; Kim et al., 2019). 도암호에서 동물플랑크톤의 현존량이 급감하는 원인은 강우 시 유입수에 의한 물리적인 씻김현상과 관련성이 있다(You et al., 2010). Beaver et al.(2013)은 Missouri River에 속하는 저수지에서 유입수량 및 방류량의 증가는 동물플랑크톤의 현존량 감소를 초래한다고 언급한 바 있다.
도암호에서는 하천으로부터의 유입수량의 강도에 따른 수체교란이 2017년에 비해 2016년과 2018년에 큰 것으로 확인되었다. 2018년의 경우, 지속적이고 빈번한 강우의 영향으로 도암호의 유입수량은 크게 변동하였다. 일반적으로 댐의 수위조절을 위해 수문을 개방하는 인공호들과는 달리, 도암호는 2001년 운영이 중단된 이후 표층방류를 통한 자연 월류식에 의존하고 있다. 따라서 동물플랑크톤의 현존량 감소의 원인은 2018년과 같이 빈번한 강우로 인한 유입수량이 많아지면서 호내 물 흐름의 형성으로 인한 물리적인 씻김의 영향이라 판단된다.
도암호에서 동물플랑크톤 군집의 계절변동은 빙어를 포함한 부유성 어류에 의한 포식압으로 이해되기 보다는 몬순강우 시 탁수유입과 체류기간의 장기화에 의한 영향 그리고 증가된 유입수류량에 의한 동물플랑크톤의 씻김현상이 복합적으로 작용하는 것으로 판단된다. 따라서 상위소비자의 포식압에 의한 영향보다는 표층방류를 통한 자연 월류를 하는 도암호의 수문학적인 특성이 동물플랑크톤 군집의 계절변화에 반영된 결과로 판단된다.
Daphnia 출현과 투명도와의 상관관계
인공호에서 수체의 투명도가 높아지는 현상을 청수기 현상(Clear water phase)이라 하며, 이는 대형동물플랑크톤인 지각류가 식물플랑크톤을 섭식 제거하는 과정에서 나타나는 현상으로 알려져 있다(Deneke and Nixdorf, 1999; Kim et al., 2003; Talling, 2003). 또한 일반적으로 몬순지역 대형인공호소에서 확인되는 청수현상은 강우 직전 수온이 증가하기 시작하는 6월∼7월과 강우 이후 호소 중층을 통해 영양염이 재공급되는 9월 중에 나타나는데, 이 기간에는 Daphnia spp.의 출현이 있고 현존량 또한 높아지는 시기이다.
도암호에서 투명도(Secchi depth; SD)가 높게 확인된 2017년 6월은 Daphnia galeata의 현존량이 급증한 시기였다. 이 시기의 투명도는 댐앞 지점(DAD1)에서 3.7 m로 확인되었고 수체가 가장 안정되고 수온성층이 발달되어 있으며 표층수온이 22°C까지 증가한 시기였다. 2017년(0.3 m-3.7 m; 1.0 ± 0.7)의 투명도는 2016년(0.7 m-1.5 m; 0.9 ± 0.2)과 2018년(0.3 m-1.2 m; 0.8 ± 0.3)의 투명도에 비해 다소 높았다. 하지만 2017년 6월 이외의 조사 시기에는 청수현상이 나타나지 않았고, 수온성층이 소멸되면서 수체 전체가 순환할 때에는 더욱 투명도가 낮아지는 양상이 매년 반복되었다. 도암호에서 청수현상은 2017년 6월에 댐앞 지점(DAD1)에서 확인되었으나 몬순강우 이후 온대호소에서 나타나는 2차 청수현상 또한 확인되지 않았다. 이는 도암호의 방류방식이 자연 월류식이고 더욱이 매년 탁수의 영향에 호내 전 수체가 반복적으로 노출되기 때문으로 판단된다.
본 연구에서 2016년과 2018년 6월에는 동물플랑크톤 군집조사를 하지 않았으나 이들 해에는 2017년 6월과 달리 낮은 투명도를 보였다(Fig. 4). 6월에 청수현상이 확인된 2017년에는 Daphnia galeata, Diaphanosoma brachyurum의 출현이 있던 반면, 2016년과 2018년의 동물플랑크톤 군집구조는 대형동물플랑크톤의 출현이 없이 소형지각류인 Bosmina longirostris, Bosminopsis deitersi의 빈번한 출현이 특징적이다. Talling(2003)에 의하면 청수현상은 소형지각류인 Bosmina spp.가 출현하는 시기에는 나타나지 않았고 Daphnia galeata가 출현하는 시기에 국한되어 나타나는 현상이라고 하였다. 또한 Olds et al.(2014)는 요각류 및 동물플랑크톤 군집의 경우 Chl-a농도 및 투명도와 양의 상관성을 보이는 반면, Daphnia spp.는 음의 상관성을 보임을 보고한 바가 있다. 이와 같은 연구 결과들은 도암호에서 2017년 6월에 국한되어 청수현상이 나타난 이유를 설명해 준다.
따라서 도암호에서 조사 기간 중 2017년 6월 이외의 시기에 대형지각류인 Daphnia galeata가 출현하지 않는 원인 중 하나로 수체의 체류시간(수체의 안정화)과 탁수발생으로 인한 요인이 크다고 판단된다. 2016년과 2018년에는 공통적으로 Daphnia galeata가 출현하는 시기를 전후로 빈번한 유입수량의 변동이 있었고 이로 인한 수체의 교란과 고탁수의 영향이 지속되었기 때문에 Daphnia galeata가 출현하기에는 부적합한 환경조건이었을 것으로 판단된다(Beaver et al., 2013; Hart, 1988). Hart(1988)는 동물플랑크톤 종에 따른 탁수의 내성이 다름을 보였는데, Daphnia 속이 Moina 속이나 요각류에 비해 탁수에 대한 내성이 낮았다. 또한, 탁수의 영향을 받는 호소에서 동물플랑크톤의 먹이섭식율은 탁도가 증가할 수록 감소함을 보였다.
본 연구의 결과, 도암호의 동물플랑크톤 군집구조는 소형동물플랑크톤인 윤충류가 우점하는 호소생태계임을 확인하였다. 도암호에서 탁수의 영향이 지속적으로 유지될 경우 동물플랑크톤 군집 내 대형동물플랑크톤의 출현 빈도는 더욱 낮아질 것이고, 소형동물플랑크톤 위주의 microbial loop에 의한 먹이망구조가 고착화될 것으로 사료된다. 따라서, 도암호에서 확인된 청수현상의 출현 빈도 및 형성 기간은 향후 동물플랑크톤 군집구조의 변화와 더불어 중요한 서식환경 변화를 이해하는 지표가 될 것으로 판단된다(Chen and Chen, 2017).
도암호는 송천 상류유역 고랭지경작지에서 발생하는 탁수의 영향을 지속적으로 받고 있는 호소생태계이지만, 동물플랑크톤 출현종 수 및 현존량의 계절변동 및 군집구조는 대부분의 부영양호의 동물플랑크톤 군집구조와 유사한 양상을 보였다.
결론적으로 도암호의 동물플랑크톤 군집에 영향을 미치는 요인은 강우패턴의 변화, 매년 반복되는 송천 상류유역 고랭지경작지에서 유입하는 탁수 그리고 수리수문학적인 특성으로 파악되었으며, 도암호는 호내에 지속적으로 체류하며 누적되는 탁수입자의 영향이 복합적으로 반영된 호소생태계로 파악되었다.
요 약
본 연구는 자연 월류식 표층방류를 하는 도암호를 대상으로 동물플랑크톤 군집구조의 시・공간적인 특성을 파악하기 위하여 2016년부터 2018년까지 총 12회 수질 및 생물분포를 조사하였다. 조사기간 동안 누적된 연도별 총 현존량은 2016년 1,942.3 ind./L(70.9 ind./L-374.1 ind./L), 2017년 2,852.6 ind./L(47.2 ind./L-906.5ind./L) 그리고 2018년 985.5 ind./L(10.4 ind./L-312.5 ind./L)로 확인되었고, 총 출현종 수는 2016년 29종(8종-19종), 2017년 34종(2종-18종) 그리고 2018년 25종(5종-15종)이었다.
연도별 동물플랑크톤 종수 및 현존량의 차이는 몬순강우 시 도암호의 수리・수문학적 특성에 의한 탁수층의 발달 및 긴 체류기간과 관련이 있는 것으로 나타났다. 도암호 내에 형성된 탁수층의 지속기간은 2016년과 2017년에는 7월과 8월 사이였고, 2018년에는 5월부터 9월까지 4개월간 지속되었다. 도암호의 동물플랑크톤 군집구조를 파악한 결과, 윤충류가 33종(68.8%)으로 가장 많았고 요각류(8종; 16.7%), 지각류(4종, 8.3%) 그리고 원생동물(3종, 6.3%) 순으로, 소형동물플랑크톤 군집구조를 보였다. 청수기는 조사 기간 중 유일하게 2017년 6월에 확인이 되었으며, 이 시기의 호내 특징은 탁수농도가 낮고 수온성층이 형성되어 있으며 수체가 안정된 시기로, Daphnia galeata가 출현한 것으로 확인되었다. 동물플랑크톤 군집구조의 변화는 강우패턴과 호내 탁수층의 지속기간 등에 의한 수리・수문학적인 요인과 관련성이 있는 것으로 판단되었다. 동물플랑크톤 군집구조는 탁수의 방류방식 및 체류기간으로 인한 영향을 반영하는 지표로써 활용될 수 있으며, 특히, 대형동물플랑크톤의 출현 유무는 수질환경의 변화를 파악함에 유용한 과학적 근거 자료로 활용될 것으로 판단된다.
