

Introduction
Materials and Methods
Study Period and Sites
Survey Methods
Results and Discussion
Environmental Characteristics Analysis: Focusing on Water Temperature, pH, Electrical Conductivity, Salinity, and Nutrients
Analysis of Phytoplankton Community and Species Occurrence Characteristics
Dominant Species Analysis and Environmental Implications
Diversity Index
Conclusion
Introduction
Wetlands are among the most vital components of global ecosystems, forming unique transitional zones where terrestrial and aquatic ecosystems converge and serve as crucial reservoirs for biodiversity. These areas provide habitats, refugia, and feeding grounds not only for aquatic organisms, such as insects, amphibians, and fish, but also for diverse terrestrial species, including insects, mammals, and birds (KNPS, 2007; Park et al., 2008). Furthermore, wetlands are key locations where the biogeochemical cycling of inorganic elements actively occurs, contributing significantly to ecosystem material circulation and water purification (Sze, 2003). As of 2020, South Korea has 2,323 wetlands, including 6,786 species and 116 endangered species, demonstrating their exceptionally high ecological value (NIE, 2022). However, 22 invasive alien species have been identified and their proliferation has increasingly underscored the importance of wetland ecosystem conservation and sustainable management.
Among the various wetland types in South Korea, lagoons represent unique aquatic systems with distinct ecological values. They are characterized by their brackish nature, where freshwater and seawater mix. Notably, 19 of the 21 lagoons in South Korea are distributed along the East Coast of Gangwon-do, representing the geomorphological and ecological characteristics of the region (NIE, 2022). Lagoons are formed by sandbars or spits separating them from the sea. The mixing of fresh and marine waters provides habitats for diverse, rare species and offers high scenic value. However, in recent decades, increased urbanization and recreational activities have led to a growing tendency to manage the lagoons along the East Coast of Gangwon-do (e.g., Songjiho, Gyeongpoho, Cheongchoho, and Yeongrang Lake), with a focus on development rather than natural conservation (Yoon et al., 2008). This approach has weakened the original ecological functions of lagoons and resulted in various environmental problems, including water quality degradation and ecosystem disruption.
Research on Korean lagoons has primarily focused on water quality characteristics and phytoplankton communities. Studies have been conducted on Hwajinpo and Yeongrang Lake (Mitamura and Cho, 1984), Songjiho and Yeongrang Lake (Moon et al., 2007), and Maeho (Um, 1998). Numerous studies have investigated the correlation between water quality and phytoplankton (Cho and Park, 1969; Heo et al., 1999; Moon et al., 2015; Shin et al., 2021). These studies have contributed to our understanding of patterns of environmental change in lagoons.
Yeongrang Lake, a lagoon located in the northern part of Sokcho-si, has long been recognized for its high scenic value. However, its lake area, which was approximately 1.11 km2 in 1918, decreased to 0.96 km2 by 1990 (Yoon et al., 2008). More recently, persistent developmental pressures, including the reclamation and construction of lakeside roads, golf courses, and resorts, have significantly altered both the landscape and ecological functions. In particular, water quality degradation due to non-point source pollution has led to frequent environmental problems, such as algal blooms (Heo et al., 1999), eutrophication (WREO, 1999), fish kills (Kim et al., 2008), deep-layer hypoxia and hydrogen sulfide generation (Huh et al., 2017), and winter red tide events (Park et al., 2022). In response, Sokcho-si implemented measures to improve the water quality, including sediment dredging (WREO 1999) and seawater inflow projects (Heo et al., 2011; Park et al., 2015). Recently in 2021, additional anthropogenic activities, such as the installation of a pontoon bridge, have also occurred. Amidst these changes, the Yeongrang Lake ecosystem is undergoing a complex transition from a freshwater/brackish environment to a marine-dominated environment; however, systematic long-term monitoring and ecological impact assessment studies on this transition remain insufficient.
Phytoplankton are primary producers in aquatic ecosystems and are highly sensitive to changes in water temperature, salinity, and nutrient levels. Community structure and succession of dominant species serve as crucial bioindicators for assessing water quality and ecosystem health (Sze, 2003). In particular, in lagoons experiencing rapid environmental changes owing to seawater inflow, changes in phytoplankton communities can sensitively reflect the overall ecosystem transition.
Therefore, this study aimed to analyze the spatiotemporal dynamics of phytoplankton communities in Yeongrang Lake, a lagoon subjected to prolonged anthropogenic interventions. By comparing these findings with those of previous studies in other lagoons, we evaluated the unique characteristics of the Yeongrang Lake ecosystem and the extent of its marine transformation. This study has significant implications for understanding ecological changes in Yeongrang Lake and provides scientific evidence for effective future environmental management and conservation strategies.
This study addresses the following research questions:
1. To what extent has Yeongrang Lake transitioned from a lagoon to a marine-dominated waterbody?
2. How have phytoplankton community composition and dominant species changed in response to environmental alterations?
3. What is the relationship between these community changes and ecosystem stability as well as the potential risk of harmful algal blooms?
Materials and Methods
Study Period and Sites
To analyze the spatiotemporal dynamics of phytoplankton communities in response to seawater intrusion in Yeongrang Lake, four seasonal surveys were conducted from December 2022 to October 2023. Five sampling stations were selected: one river inflow point (R; River) and four stations within Yeongrang Lake (St.1-4) (Table 1, Fig. 1). At each station, both surface and bottom layer samples were collected. The bottom-layer samples were collected approximately 1 m above the sediment, considering the water depth at each sampling time. The linear distance between the river inflow point (R) and Yeongrang Lake St.1 was approximately 212 m, and the average distance between stations St.1 and St.4 within the Yeongrang Lake was approximately 400 m. All sampling stations were located 0 m above sea level. The river inflow station (R) was included to identify the characteristics of freshwater-origin phytoplankton input and evaluate the freshwater-seawater mixing status within Yeongrang Lake.
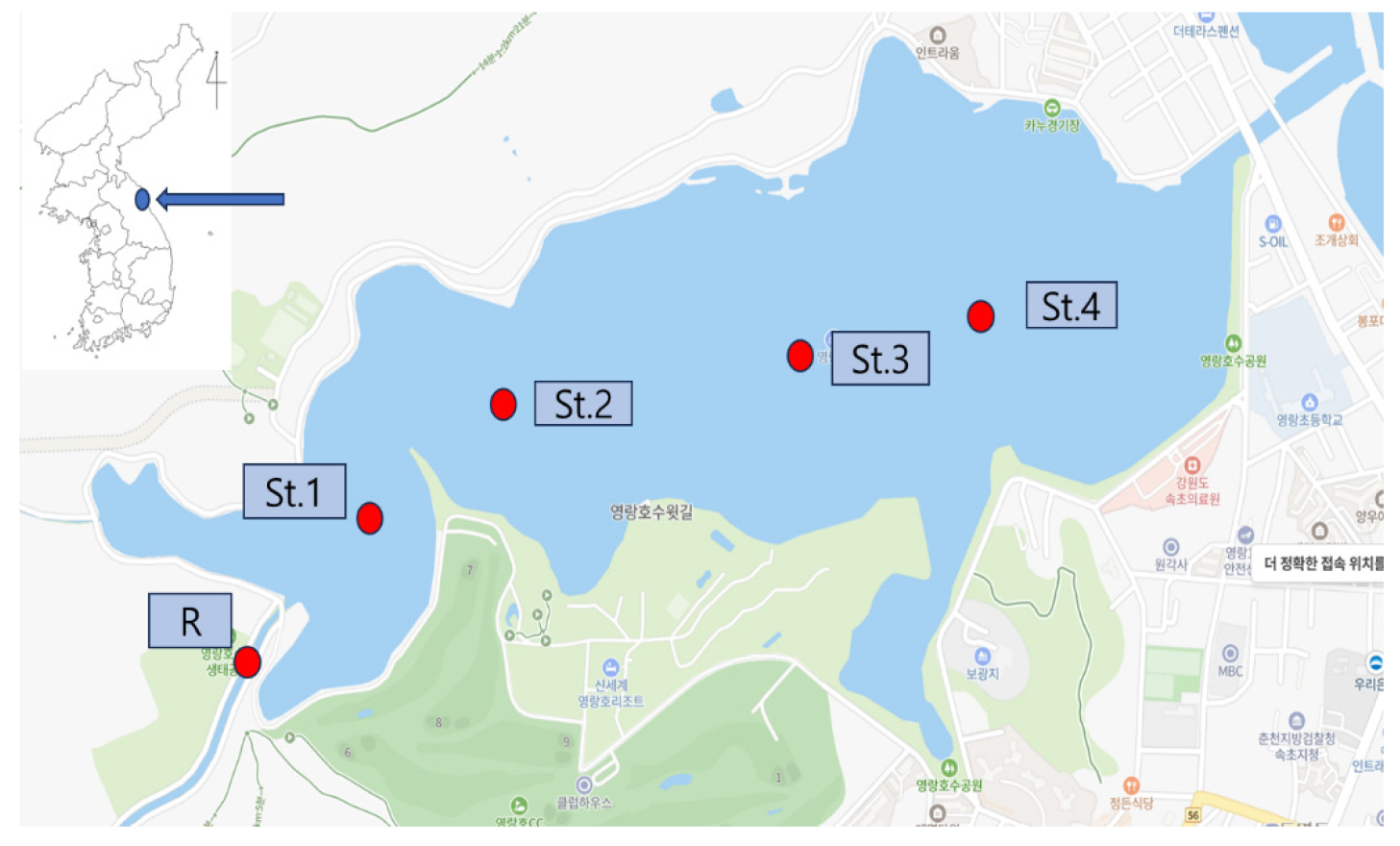
Table 1.
Survey Period and Geographic Coordinates of Survey Stations in Yeongrang Lake
| Survey period | Sampling sites | Coordinate | |
| Winter : 2022.12.28–12.29 | R* | 38° 12′ 49.58″ | 128° 34′ 12.08″ |
| Spring : 2023.03.29–03.30 | St.1 | 38° 12′ 53.50″ | 128° 24′ 21.26″ |
| Summer : 2023.07.11–07.12 | St.2 | 38° 13′ 02.36″ | 128° 34′ 29.95″ |
| Fall : 2023.10.04–10.05 | St.3 | 38° 13′ 04.06″ | 128° 34′ 44.61″ |
| - | St.4 | 38° 13′ 06.33″ | 128° 35′ 04.70″ |
Survey Methods
To analyze the habitat and community characteristics of phytoplankton, surveys were conducted using the following methods.
Environmental Factor Measurement
Environmental factors, such as water temperature, pH, electrical conductivity (EC), and salinity, were measured directly in the field using a portable meter (YK-2001PHA, Lutron). Nutrient concentrations, specifically total nitrogen (T-N) and total phosphorus (TP), were analyzed using public data from the National Institute of Environmental Research’s Water Environment Information System (NIER, n.d., http://water.nier.go.kr).
Sample Collection and Pretreatment
For the qualitative analysis of phytoplankton communities, samples were collected from the surface water using a plankton net (Rhigosha Nxxx25, ø20 µm). For quantitative analysis, surface samples were collected using 1 L polyethylene bottles at a depth of 10-30 cm. Bottom-layer samples were collected using a Van Dorn water sampler (1.5 L) at a designated depth (1 m above the bottom) at each station. Immediately upon collection, 3 mL Lugol’s solution was added to fix the samples, and an additional 0.5-1 mL of formalin solution was added for long-term preservation.
Quantitative Analysis and Identification
For quantitative analysis, the samples were allowed to settle for 48-72 h, after which the supernatant was removed using a siphon to achieve a final concentration of 100 mL. Subsequently, 1 mL of the concentrated sample was introduced into a S-R chamber (Sedgwick-Rafter chamber) and counted under a microscope to calculate the phytoplankton density (cells/mL).
As diatom identification relies on rigid siliceous valves, a separate cleaning process is required. Diatom samples were cleaned using hydrochloric acid, sulfuric acid, and potassium permanganate. Permanent slides were prepared using mounting media (Fujifilm). The identification was performed using an optical microscope.
Species Identification Criteria
For phytoplankton species identification, the following taxonomic guides were referenced:
∙ Chlorophyta: Komárek and Fott (1983), Kanetsuma and Yamagishi (2018)
∙ Cyanobacteria: Komárek and Anagnostidis (2001)
∙ Freshwater benthic diatoms: Cantonati et al. (2017), Watanabe (2005)
∙ Marine phytoplankton: Hoppenrath et al. (2009), Kraberg et al. (2010), Witkowski et al. (2000)
The taxonomic system of the identified species was organized based on Guiry and Guiry’s (2023) AlgaeBase (https://www.algaebase.org).
Results and Discussion
Environmental Characteristics Analysis: Focusing on Water Temperature, pH, Electrical Conductivity, Salinity, and Nutrients
In this study, the key environmental factors of Yeongrang Lake were measured and analyzed to gain an in-depth understanding of the characteristics of lake ecosystems.
Water Temperature Changes
The water temperature in Yeongrang Lake exhibited similar trends in both surface and bottom layers. Surface water temperatures varied widely from 2.7°C (St. 3 in winter) to 27.1°C (St. 4 in summer), with an average temperature of 19.3°C. The bottom layer also ranged from 3.4°C (St. 4 in winter) to 26.6°C (St. 2 in summer), recording an average of 19.2°C (Table 2). Spatially, the average water temperature in the upstream area of the pontoon bridge (St.1, St.2) was approximately 21.8-22.1°C, which was about 4.5-4.8°C higher than the average temperature in the downstream area (St. 3, St. 4) (17.0-17.6°C). This suggests that the water flow stagnation in the upstream pontoon bridge area likely led to increased water temperatures. The overall average water temperature for Yeongrang Lake was 19.3°C, while the river inflow point (R) showed an average of 16.9°C (ranging from 5.9-25.9°C).
Compared to previous studies (Bhattrai et al., 2017), the average water temperature of Yeongrang Lake in this study was approximately 5.1°C higher than the average of 14.2°C recorded from 1998 to 2015. Furthermore, compared to the survey conducted from April to October 2011 (Moon et al., 2015), the average water temperature during the same period in this study was about 3.9°C higher. This indicates a continuous increase in the water temperature of Yeongrang Lake, aligning with global temperature increases or suggesting the involvement of factors directly or indirectly influencing water temperature increases (e.g., decreased inflow volume and degree of openness of inflow rivers). Such increases in water temperature are closely related to the physiological activity of aquatic organisms (Lee and Han, 2007) and are expected to significantly influence the species composition, community structure, and dominant phytoplankton species in Yeongrang Lake in the future.
pH Changes
The pH of the surface layer of Yeongrang Lake ranged from 7.95 (St. 1, autumn) to 8.14 (St. 2, autumn) (Table 2). The pH of the bottom layer varied from 7.74 (St. 2; autumn) to 8.19 (St. 3; winter). At most stations, the surface pH was slightly higher than that of the bottom layer, except at St. 4, where the bottom layer pH was higher than that of the surface. The pH at the river inflow point (R) fluctuated widely, ranging from 6.46 to 7.97.
Electrical Conductivity (EC) and Salinity
The EC of the surface layer ranged from 42.6 mS/cm (St.1, summer) to 51 mS/cm, with an average of 47.62 mS/cm. The EC values of the bottom layer recorded EC values from 46 (St.4, winter) to 52 mS/cm, with an average of 49.84 mS/cm (Table 2). The average EC of the bottom layer was slightly higher than that of the surface layer. Salinity in the surface layer ranged from 27.6 psu (St. 3, autumn) to 30.5 psu (St. 2, spring) with an average of 28.8 psu. The bottom layer exhibited salinity values from 27.9 psu (St. 4, autumn) to 32.1 psu (St. 4, spring), averaging 29.7 psu (Table 2). Similar to the EC, the average salinity of the bottom layer was slightly higher than that of the surface layer. In contrast to a previous study in 2015 (Moon et al., 2015) where Yeongrang Lake’s salinity was 7.8 psu, exhibiting characteristics of a typical lagoon, this study observed a significant increase in salinity, confirming the ongoing marine transformation of the lake. This is presumed to be the result of artificial efforts, including sediment dredging for water quality improvement (WREO, 1999) and securing seawater inflow channels to address bottom-layer hydrogen sulfide accumulation (Hong et al., 2024).
Nutrient Concentrations (Total Nitrogen [TN] and Total Phosphorus [TP])
The TN concentrations ranged from 0.164 to 0.458 mg/L, with an average of 0.323 mg/L (Table 2). Concentrations were high in May and June, decreased in July, increased again in August, and approached a maximum in October. Past studies have shown a continuous decreasing trend in T-N concentration, with values of 2.13 mg/L in 1999 (Heo et al., 1999) and 0.502 mg/L in 2017 (Huh et al., 2017). It is presumed that the inflow of nitrate substances around Yeongnangho Lake has been significantly improved.
TP concentrations ranged from 0.015 to 0.048 mg/L, with an average of 0.029 mg/L; the highest concentration was recorded in August (Table 2). The T-P concentration also showed a gradual decrease, similar to T-N, considering the values of 0.446 mg/L in 1999 (Heo et al., 1999) and 0.059 mg/L in 2017 (Huh et al., 2017).
Meanwhile, the T-N/T-P ratio ranged from 10.92 to 62.29, with an average of 33.93 (Table 2). It was the highest in May and June (57.25-62.29), sharply decreased in July, and then increased again from August, approaching the maximum value of 60 in October. Huh et al. (2017) reported a strong inverse correlation between the increase/decrease in TN and TP concentrations and salinity in Yeongrang Lake. Furthermore, considering that the water quality of the Jangcheon River, the largest inflow into Yeongrang Lake, is classified as Class II (slightly good) according to river water standards, and acknowledging existing reports suggesting its minimal impact on Lake’s water quality improvement (Huh et al., 2017), it is inferred that efforts to introduce seawater contribute to the reduction of T-N and T-P.
Considering the Redfield ratio (TN/TP = 16:1) for optimal algal growth (Redfield, 1934), the T-N/T-P ratio in this study was lower than that of artificial lakes (40-100) but higher than that of other lagoons (Cheongchoho 17.0-40.5, Maeho 4.0-18.8, Hyangho 7.7-19.0) (Kang and Choi, 2002; Kim et al., 1997; Moon et al., 2015). This suggests that nitrate substances entered Yeongrang Lake from external sources. Although phosphate appears to be a limiting factor for phytoplankton growth in Yeongrang Lake, more so than nitrate, it remains unclear whether nutrient deficiency is a direct limiting factor for phytoplankton growth, given the generally high concentrations of both nitrogen and phosphorus (Heo et al., 1999). This analysis of environmental factors provides a crucial basis for understanding the ecological characteristics and changing patterns of Yeongrang Lake and can serve as foundational data for future sustainable management.
Table 2.
Fluctuations of environmental factors at the study sites in Yeongrang Lake
Analysis of Phytoplankton Community and Species Occurrence Characteristics
This study conducted an in-depth analysis of phytoplankton community characteristics identified in Yeongrang Lake and its inflowing river (R).
Total Phytoplankton Species Occurrence and Taxonomic Composition
In total, 163 phytoplankton species were identified, encompassing both Yeongrang Lake and the inflowing river (R). Among these, diatoms showed the highest occurrence with 100 species (61.3%), followed by Chlorophyta with 35 species (21.5%), Dinophyta with 15 species (9.2%), Euglenophyta with 8 species (5%), Cryptophyta with 3 species (1.8%), and Charophyta and Heterokontophyta with 1 species each (0.6%).
Phytoplankton Community Characteristics in Yeongrang Lake and the River
A total of 99 species were identified in the Yeongrang Lake. Of these, 77 species (74.8%) were diatoms and 15 species (24.26%) were dinoflagellates, making these two taxonomic groups the most dominant in Yeongrang Lake. Additionally, two species each of Chlorophyta and Euglenophyta and three species of Cryptophyta were identified (Fig. 2). Similar to the general phenomenon observed in Korean coastal waters, where diatoms and dinoflagellates show high proportions (Choa and Lee, 2000; Park et al., 2015), Yeongrang Lake also exhibited the dominance of these two groups. Notably, unlike the 2011 study (Moon et al., 2015) where cyanobacteria showed a high proportion of diatoms, no cyanobacteria were observed in this study. This suggests that the phytoplankton community of Yeongrang Lake has shifted towards marine characteristics due to water quality improvement efforts, such as sediment dredging (WREO, 1999) and the artificial opening of channels for seawater inflow (Huh et al., 2017).
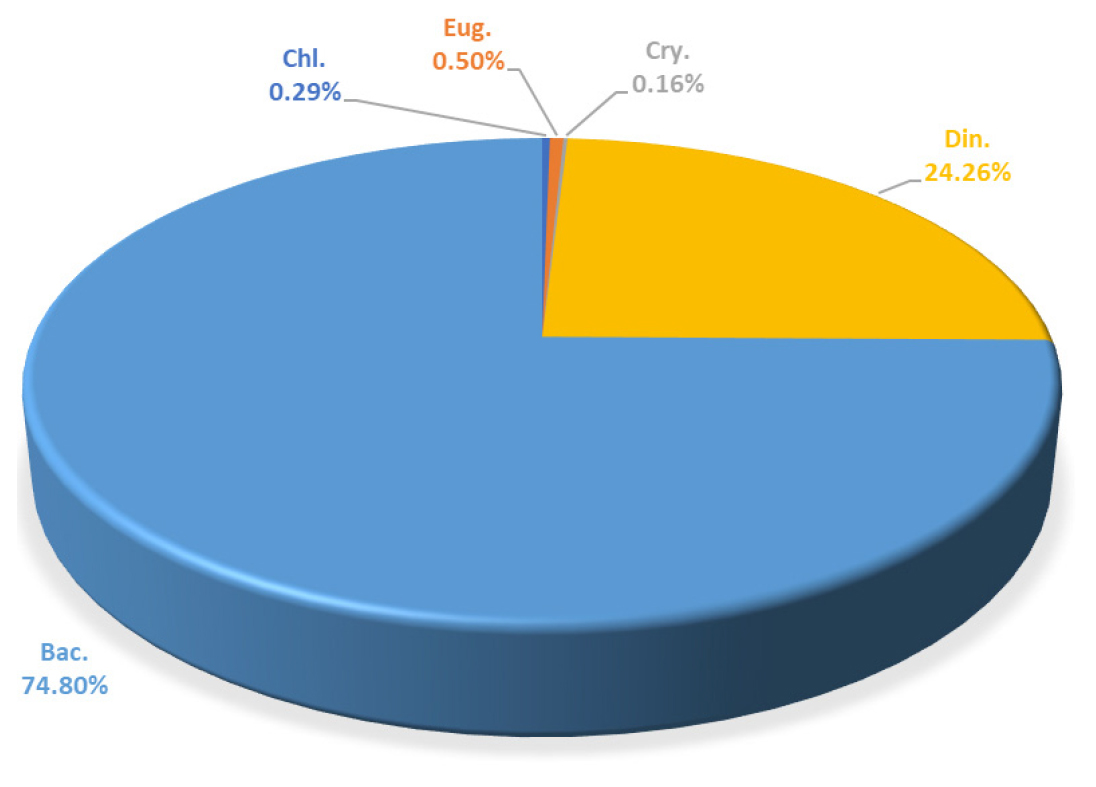
Seventy-one phytoplankton species were identified in the river (Station R), with Chlorophyta (33 species) and diatoms (30 species) being the dominant taxonomic groups. Euglenophyta (six species), Charophyta, and Heterokontophyta (one species) were also observed. Interestingly, dinoflagellates and cryptophytes were found exclusively in Yeongrang Lake, while Charophyta and Heterokontophyta were found only in the river, clearly demonstrating distinct taxonomic compositions based on the habitat environment.
Number of Species Occurring by Station and Season
The total number of species observed per sampling station was the highest at St. 1 and St. 4 (52 species each) and the lowest at St.2 (47 species). The river station (R) contains 71 species. Seasonally, species occurrence ranged from 4 to 18 species. In the surface layer, the highest number of species (14) was observed in autumn, whereas in the bottom layer, the highest number (17) was observed in spring. The river station had the highest number of species (49) in the spring.
Major Dominant Genera and Common Species Between River and Lake
The diatom Chaetoceros (13 species) was the most frequently observed genus in Yeongrang Lake, followed by Nitzschia (12 species), Navicula and Gyrosigma (seven species each), the dinoflagellate Prorocentrum (six species), Protoperidinium, and the diatom Rhizosolenia (five species each). In the river, the Chlorophyta genus Scenedesmus (15 species) was the most dominant, followed by Navicula (six species), Monoraphidium, Aulacoseira, and Nitzschia (four species each).
Ten species were common between the rivers and Yeongrang Lake. These included two Chlorophyta species (Dictyosphaerium indicum and Pediastrum duplex) and eight diatom species (Aulacoseira granulata, Aulacoseira granulata var. angustissima, Cyclotella meneghiniana, Gyrosigma acuminatum, Melosira varians, Navicula gregaria, Nitzschia acicularis, and Synedra ulna). These species are commonly found in freshwater environments. N. gregaria in particular, is a diatom species that inhabits both freshwater and brackish waters and is frequently observed in highly organically polluted freshwater areas.
Comparison of Species Occurrence Between Upstream and Downstream of the Pontoon Bridge
A comparison of the phytoplankton species occurrence between the upstream (St.1, St.2) and downstream (St.3, St.4) areas of the pontoon bridge revealed 42 common species (2 Euglenophyta, 2 Cryptophyta, 10 Dinophyta, and 28 diatoms). The upstream areas recorded 32 species (including 2 Chlorophyta and 30 diatoms), whereas the downstream areas recorded 25 species (including 1 Cryptophyta, 5 Dinophyta, and 19 diatoms). Although seven more species were found in the upstream areas, the phytoplankton were microscopic and moved with water currents, making it difficult to definitively determine the species differences caused by the pontoon bridge based solely on short-term surveys. Therefore, long-term monitoring and data collection are required.
Overall, over 95% of the 99 species identified in Yeongrang Lake were marine, suggesting that Yeongrang Lake is gradually transitioning into a marine water body rather than functioning as a mixed freshwater-seawater lagoon. Meanwhile, differences in water quality, such as the influence of water flow due to the pontoon bridge, appeared to have caused site-specific variations in the number of phytoplankton species within the lake.
Total Phytoplankton Abundance and Taxonomic Density
The total phytoplankton abundance in Yeongrang Lake ranged from 3,234 to 6,139 cells/mL in both the surface and bottom layers. Abundance was higher in the bottom layer at St. 1-St. 3, but slightly higher in the surface layer at St. 4.
Regarding taxonomic abundance, Chlorophyta, Euglenophyta, and Cryptophyta showed relatively low densities, ranging from 3 to 107 cells/mL, and were often completely absent at certain stations or seasons (Table 3). Dinoflagellates were more abundant in the bottom layer, ranging from 3 to 840 cells/mL in the surface layer and 3 to 2,281 cells/mL in the bottom layer (Fig. 3). Seasonally, the highest abundance of dinoflagellates was observed in summer in both surface and bottom layers. Diatoms ranged from 3 to 4,823 cells/mL in the surface layer and from 3 to 4,710 cells/mL in the bottom layer, showing a tendency to be higher in spring and autumn, with the maximum abundance observed in autumn.
Table 3.
Yearly changes in dominant phytoplankton species in Yeongrang Lake
| Years | Dominant species | Month | Reference |
| 1999 | Microcystis spp. Oscillatoria spp. | August | Heo et al. (1999) |
| 2009 | Cryptomonas cf. marssonii Heterocapsa rotundata | January | Park et al. (2022) |
| 2011 | Cyclotella striata Skeletonema costatum Euglena gigans |
April, August June October | Moon et al. (2015) |
| 2023 | Chaetoceros decipiens Cylindrotheca closterium Navicula gregaria Prorocentrum minimum Prorocentrum redfieldii |
March Allseason January March August | This Year |
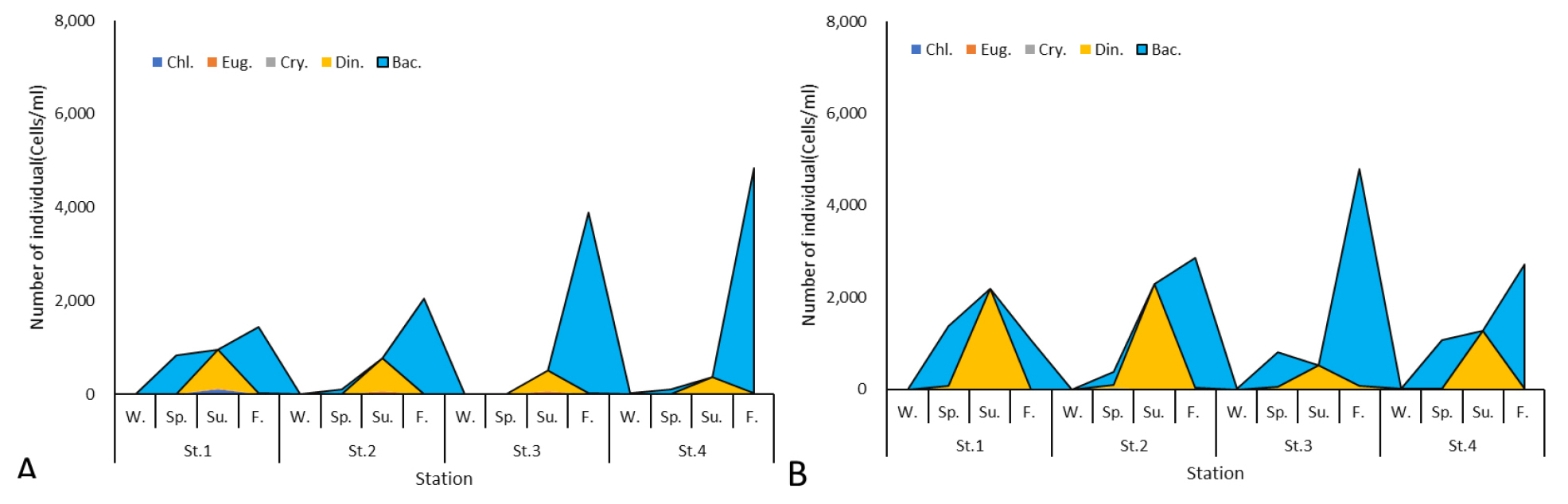
The total abundance in the upstream pontoon bridge area was 4,644 cells/mL at St.1 and 5,527 cells/mL at St.2. In the downstream area, St.3 recorded 6,139 cells/mL and St.4 recorded 5,092 cells/mL, indicating a slightly higher total abundance in the central part of the Yeongrang Lake survey area.
Dominant Species Analysis and Environmental Implications
This study identified five dominant phytoplankton species in Yeongrang Lake: three diatom species (Chaetoceros decipiens, Cylindrotheca closterium, N. gregaria) and two dinoflagellate species (Prorocentrum minimum and Prorocentrum redfieldii). C. decipiens was excluded from the main analysis because of its temporary occurrence, with a density of only 150 cells/mL in the surface layer of St.2 during spring.
C. closterium
C. closterium exhibited cell density fluctuations ranging from 3 to 4,640 cells/mL. High densities were observed in autumn in both the surface and bottom layers at all stations. In the other seasons, its occurrence was minimal or less than 8 cells/mL (Fig. 4). During its dominance, the average water temperature was 23.3°C, pH was 7.87, electrical conductivity was 47.6 mS/cm, and salinity was 28.2 psu. C. closterium is a common species found in most Korean coastal waters and often forms blooms, primarily in summer (Lee and Han, 2007) or autumn (Choa and Lee, 2000). Lee and Han (2007) reported that C. closterium blooms during periods of low salinity. This aligns with our findings, as the autumn salinity in this study (28.18 psu) was lower than that in the spring (average of 30.49 psu) and summer (28.75 psu).
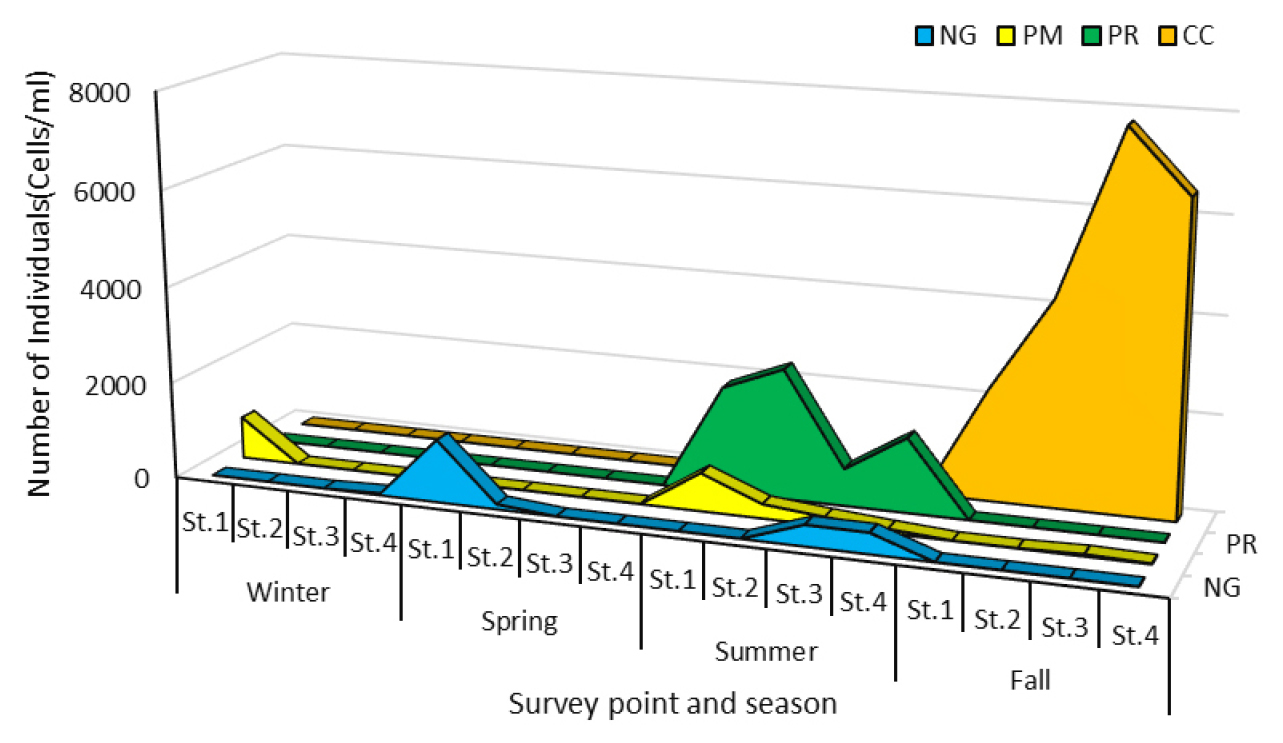
N. gregaria
N. gregaria had cell densities ranging from 40 to 862 cells/mL, with the highest counts observed in the bottom layer at St.1. High densities were recorded in both the surface and bottom layers of St.1, St.2, and St.4 during spring and in the bottom layer of St.3 and St.4 during summer. However, it was completely absent in the winter and autumn.
∙ During spring bloom: Water temperature 15.8-16.2°C, pH 7.98-8.06, electrical conductivity 51-52 mS/cm, salinity 30.3-30.7 psu.
∙ During summer bloom: Water temperature 25.8-27.1°C, pH 7.97-8.11, electrical conductivity 46-51 mS/cm, salinity 27.7 psu.
N. gregaria is a diatom species that inhabits freshwater and brackish environments, with a broad ecological range spanning from clear to polluted waters (Watanabe, 2005), and exhibits strong tolerance to wastewater (Cantonati et al., 2017). In Korea, this species is found nationwide, including in the Namhan River and Chungju Lake and is classified as an indicator of eutrophication (NNIBR, 2017). This species showed two peaks in cell density during spring and summer. The differences in environmental factors between these two periods (spring: lower water temperature and relatively higher salinity; summer: higher water temperature and lower salinity) suggest that N. gregaria possesses multiple ecotypes that have adapted to various environmental conditions (NNIBR, 2017).
P. minimum
P. minimum was observed at St.1 in winter and at St.1, St.2, and St.3 in summer and autumn. A maximum cell density of 782 cells/mL was observed in the surface layer of St.1 during the winter. At its peak abundance, the water temperature was 26.5°C, pH 8.07, electrical conductivity 42.6 mS/cm, and salinity 29.8 psu. P. minimum is known to cause red tides through mass occurrences in areas, such as Masan Bay and Jinhae Bay, during the 1980s (Choa and Lee, 2000; Lee and Han, 2007). While this species’ mass occurrence timing varies, with some studies showing it in late autumn to winter (Lee and Han, 2007) and others in spring or spring to summer (Choi et al., 2000), Yeongrang Lake exhibited unique peak abundances in both winter and summer. This is likely attributable to P. minimum’s euryhaline characteristic (optimal growth salinity 3-34 psu) and eurythermal characteristic (optimal growth temperature 3-30°C) (Iwasaki, 1983; Kunio et al., 1990), allowing it to proliferate in both low-temperature winter and high-temperature summer conditions.
P. redfieldii
P. redfieldii was observed at cell densities ranging from 8 to 2.225 cells/mL at all stations during the summer. Their abundance declined sharply in autumn, indicating that summer was the primary period of occurrence. At its peak abundance, the water temperature was 26.6°C, pH 7.87-7.92, electrical conductivity 51 mS/cm, and salinity 29.8 psu. P. redfieldii was first discovered in the brackish waters of the Woods Hole area in Massachusetts, USA; however, its geographical distribution has not yet been clearly defined (Hoppenrath et al., 2009). In the North Sea, it occurs between May and November with a peak occurring from September to November, which differs seasonally from the peak abundance observed in July in this study.
Annual Changes in Dominant Species and Environmental Implications
The dominant species in Yeongrang Lake exhibited significant annual variations (Table 3).
∙ 1999: Cyanobacteria, specifically Microcystis spp. and Oscillatoria spp., dominated and caused severe algal blooms (Heo et al., 1999).
∙ 2009: In January, during low water temperatures, the cryptophyte Cryptomonas cf. marssonii and the dinoflagellate Heterocapsa rotundata became dominant, resulting in an unusual winter red tide event (Park et al., 2022). Park et al. (2022) found that this winter red tide occurred because of eutrophication, low predation pressure, and the fulfillment of environmental conditions for cold-adapted species.
∙ 2011: Diatoms, such as Cyclotella striata, Skeletonema costatum, and the euglenophyte Euglena gigans, were dominant (Moon et al., 2015). S. costatum is commonly found in most Korean coastal and brackish waters and possesses high ecological adaptability, often forming masses regardless of the season (Kim et al., 2012).
Thus, the shifts in dominant phytoplankton communities in Yeongrang Lake followed a pattern of Cyanobacteria → Cryptophyta/Dinophyta → Diatoms/Euglenophyta → Diatoms/Dinophyta. Furthermore, it was rare for the same dominant species to persist annually or redominate during the new survey periods, indicating that Yeongrang Lake is experiencing diverse internal environmental changes. These changes in the dominant species are considered to be closely related to alterations in the water quality of Yeongrang Lake, particularly the ongoing marine transformation due to increased seawater inflow.
Diversity Index
The species diversity index serves as a crucial metric for evaluating the phytoplankton community structure, as it reflects the integrated response of the community to environmental factors (Mukai and Takimoto, 1985). Although limitations exist, such as potential counting errors stemming from a lack of specialized taxonomic expertise or uneven sample distribution within counting chambers, which can complicate the precise discernment of community differences (Duarte et al., 1990), it remains a valuable ecological indicator because it elucidates the relationship between environmental factors and phytoplankton communities (Kang and Choi, 2002).
Current Status of Species Diversity Index in Yeongrang Lake
In Yeongrang Lake, the species diversity index ranged from 0.235 to 2.084 (average: 1.205) in the surface layer and from 0.112 to 2.690 in the bottom layer. There was a general trend of higher diversity index values in spring and lower values in summer (Fig. 5).
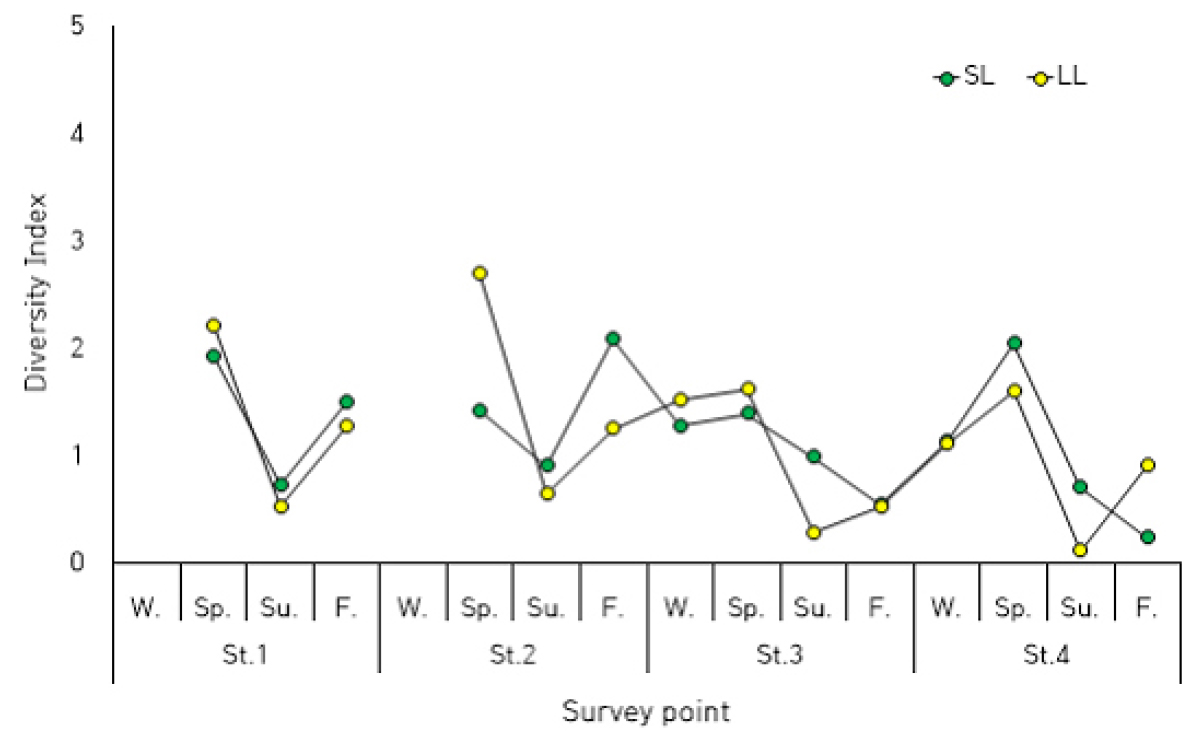
Specifically, the periods exhibiting the lowest diversity index values were as follows:
∙ Surface layer: In autumn at St.4, the diatom Cylindrotheca closterium reached an exceptionally high abundance of 4,640 cells·mL-1, with an occurrence rate of 96.2%, indicating a phenomenon of hyper-dominance.
∙ Bottom layer: In summer at St.4, the dinoflagellate Prorocentrum redfieldii had an abundance of 1,263 cells·mL-1, and its occurrence rate reached 98%.
These instances of low diversity index values strongly suggest that the community structure during these periods was highly unstable and characterized by the overwhelming dominance of a single species.
Comparison with Other Korean Coastal Waters
When comparing the fluctuation range of the Yeongrang Lake species diversity index with that of other Korean coastal areas, the following trends were observed.
∙ Jeju Island coastal waters: 0.45-2.78 (Choa and Lee, 2000)
∙ Uljin coastal waters: 1.21-2.86 (Kang and Choi, 2002)
∙ Gori coastal water: 1.05-3.14 (Kang and Choi, 2002)
∙ Gori coastal waters: 1.00-3.19 (Kang and Choi, 2002)
∙ Byeonsanbando coastal waters: 0.74-1.64 (Park et al., 2015)
The range of diversity index variation in Yeongrang Lake was somewhat higher than that observed in Byeonsanbando coastal waters. However, its range of fluctuation was smaller than that of other coastal areas, indicating a relatively stable community structure within Yeongrang Lake.
Conclusion
This study unequivocally confirmed that Yeongrang Lake has lost its lagoon characteristics and transitioned into a marine-dominated waterbody. This transformation is evidenced by rising water temperatures, increased salinity, reduced concentrations of TN and TP, and the dominance of marine phytoplankton communities. The disappearance of cyanobacteria and the prevalence of diatoms and dinoflagellates distinctly reflect this change. In the future, there is the potential for certain dominant marine species to cause red tide events, and the long-term ecological impacts of localized anthropogenic interventions (e.g., pontoon bridge installation) must be carefully monitored. Furthermore, as this ecological transition may have long-term effects on the ecosystem services and biodiversity of Yeongrang Lake, continuous and systematic monitoring and research are essential to maintain ecosystem stability and establish strategies for managing potential ecological risks.
