

Introduction
Materials and Methods
Preparation of KPB
Cell Culture
Cell viability
Measurement of Reactive Oxygen Species (ROS)
Determination of Interleukin-8 (IL-8) Secretion
Experimental Animals
Assessment of Disease Activity Index (DAI)
Histological Analysis of Colon Tissue
Measurement of IL-6 and IL-22 Levels
Statistical Analysis
Results and Discussion
Effect of KPB on Cell Viability
Inhibitory Effect of KPB on ROS Production
Inhibitory Effect of KPB on IL-8 Production
Effect of KPB on Disease Activity Index (DAI)
Effects of KPB on DSS Intake, Spleen Weight, Colon Weight, and Colon Length
Histopathological Analysis of Colon Tissue
Effects of KPB on IL-6 and IL-22 levels
Summary
Introduction
Carex pumila Thunb., a member of the Cyperaceae family, is indigenous to sandy coastal regions. Its principal constituents include several stilbene-based phenolic compounds such as (-)-ε-viniferin, miyabenol A and C, kobophenol B (KPB), and trans-resveratrol (Dávid et al., 2021). Of particular interest is KPB, a representative resveratrol oligomer characterized by a C6-C2-C6 skeleton, which has attracted research interest due to its structural complexity and potential biological activities (Keylor et al., 2015). Stilbenes and their derivatives are widely recognized for their broad spectrum of physiological effects, including anti-inflammatory, anti-cancer, anti-diabetic, cardiovascular-protective, and anti-aging properties (Baur and Sinclair, 2006; Xue et al., 2014). These attributes suggest considerable pharmacological potential for the development of natural product-based therapeutics. Nevertheless, studies on the chemical composition and pharmacological properties of Cyperaceae species remain significantly underexplored compared with ecological and taxonomic investigations. Specifically, studies evaluating the pharmacological efficacy of Carex pumila Thunb. as a coastal ecosystem resource remain extremely limited.
Resveratrol, one of the most extensively investigated stilbene compounds exhibits low bioavailability due to rapid metabolism and degradation by the gut microbiota (Willenberg et al., 2015). Notably, its physiological activity may be enhanced via complex formation with carrier proteins (Pantusa et al., 2014; Rezende et al., 2020), thus increasing scholarly interest in resveratrol oligomers. Moreover, stilbene oligomers reportedly regulate inflammation-associated signaling pathways—including NF-κB, MAPK, the NLRP3 inflammasome, and oxidative stress signaling—mechanisms closely related to the pathogenesis and progression of intestinal inflammation (Tong et al., 2020). Accordingly, KPB is expected to possess therapeutic potential for ameliorating intestinal inflammation due to structural similarity to resveratrol oligomers. However, studies investigating the anti-inflammatory effects of KPB and its role in alleviating intestinal disorders remain scarce.
Inflammatory bowel disease (IBD) is a chronic and recurrent inflammatory disorder, principally classified into ulcerative colitis and Crohn’s disease (Jairath and Feagan, 2020). Major contributing mechanisms include gut microbiota dysbiosis, immune-mediated inflammatory responses, oxidative stress, and disruption of intestinal mucosal barrier integrity (Goyal et al., 2014). Ulcerative colitis, a refractory disease characterized by inflammation restricted to the mucosa and submucosa of the colon, clinically manifests as bloody stools, diarrhea, abdominal pain, and weight loss (Conrad et al., 2014). The dextran sulfate sodium (DSS)-induced colitis animal model closely mimics the histopathological and clinical features of ulcerative colitis, making it a widely used experimental model for evaluating anti-inflammatory agents and therapeutic candidates (Chassaing et al., 2014).
Recently, natural product-based IBD therapeutics have emerged as promising next-generation treatment modalities, owing to their capacity to regulate multiple pathways (Yeshi et al., 2020). Stilbene-derived compounds have shown potential in mitigating IBD through antioxidant and anti-inflammatory mechanisms. Nonetheless, few studies have examined the biological activity of KPB, particularly its role in regulating intestinal inflammation.
The present study aimed to investigate the anti-inflammatory effects of KPB isolated from C. pumila Thunb. using an HT-29 intestinal epithelial cell model and a DSS-induced acute colitis mouse model. Furthermore, its therapeutic potential for alleviating IBD was evaluated by assessing clinical parameters, histopathological changes, and inflammatory cytokine expression.
Materials and Methods
Preparation of KPB
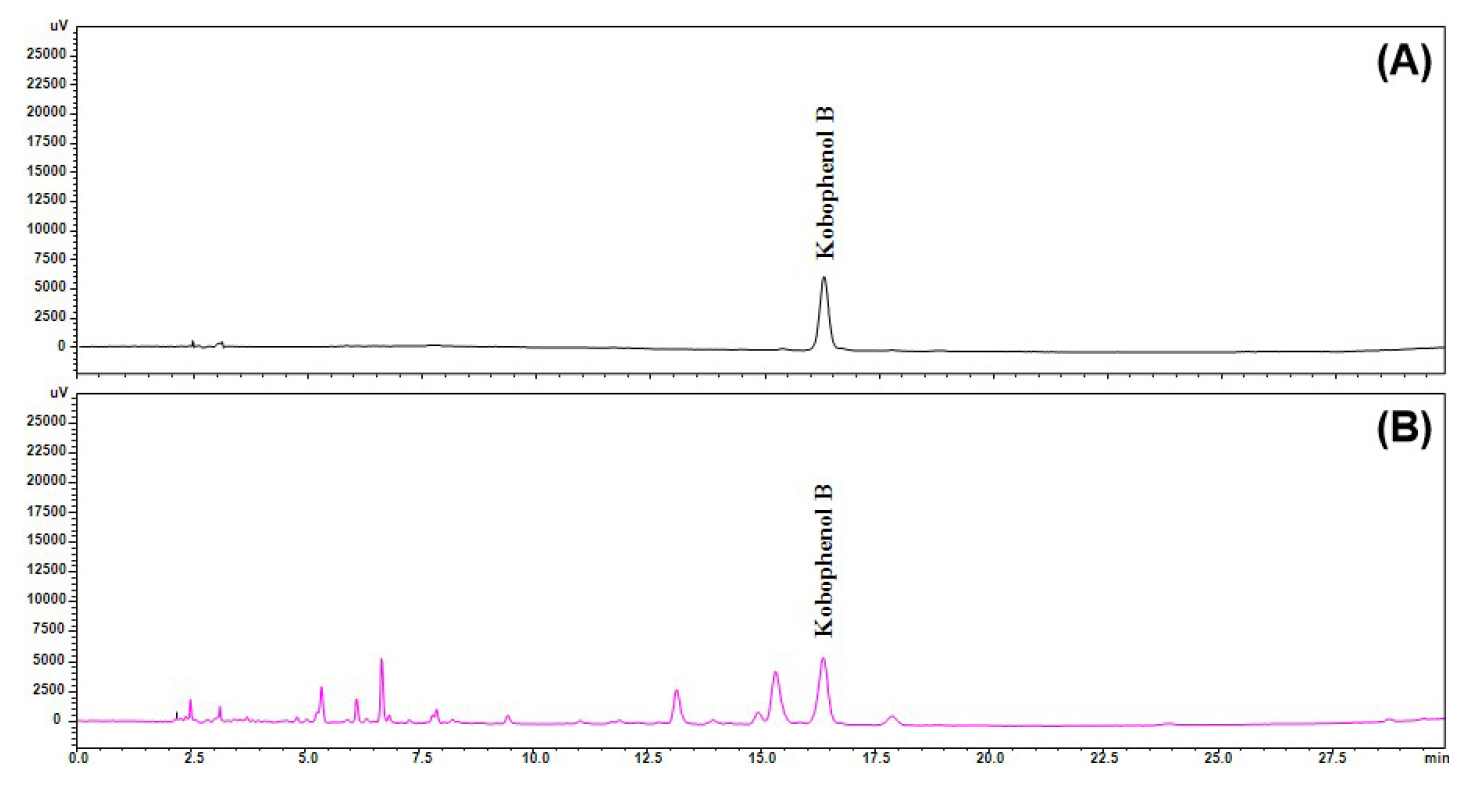
KPB was extracted from C. pumila Thunb. using 80% ethanol, and the extract was subsequently concentrated under reduced pressure. The composition of the extract was verified by high-performance liquid chromatography (HPLC) under the conditions described in Table 1 (Fig. 1). According to Korean Patent No. 10-2023-0190069, the 80% ethanol extract was fractionated stepwise using organic solvents, yielding an ethyl acetate fraction, which was purified via preparative HPLC, resulting in the isolation of KPB as a single compound. The purity of KPB was confirmed by HPLC, and its chemical structure was identified using mass spectrometry (MS) and nuclear magnetic resonance (NMR) spectroscopy (Park et al., 2025).
Table 1
Optimal HPLC-PDA analytical conditions for quercitrin in Carex pumila Thunb.
Cell Culture
The HT-29 human colorectal cancer cell line was sourced from the Korean Cell Link Bank (Seoul, Korea) and cultured in Roswell Park Memorial Institute (RPMI)-1640 medium (Corning, USA) supplemented with 10% fetal bovine serum (FBS), penicillin (100 unit/mL) and streptomycin (100 µg/mL) at 37°C in a 5% CO2 incubator (Thermo Fisher Scientific, USA). The medium was replaced every 3 days.
Cell viability
The water-soluble tetrazolium salt (WST) assay was performed to evaluate the effect of KPB on the viability of HT-29 cells. Cells were seeded at 1 × 105 cells/well in a 48-well plate and incubated for 24 h. Subsequently, the cells were treated with various concentrations of KPB (0-1600 µg/mL) and incubated for an additional 24 h. The EZ-CYTOX kit (DoGenBio, Seoul, Korea) reagent was added to each well and incubated for 1 h. The absorbance was measured at 450 nm using a microplate reader (Bio-Rad Laboratories, USA).
Measurement of Reactive Oxygen Species (ROS)
HT-29 cells were seeded at 5 × 104 cells/well in a 24-well plate and cultured for 24 h and then treated with 2% DSS and KPB (200 µg/mL) for an additional 24 h. The cells were incubated with 5 µM 2’, 7’-dichlorofluorescein diacetate (DCFH-DA) (Sigma-Aldrich, USA) at 37 °C for 45 min. After washing with PBS, fluorescence intensity was measured using a fluorescence microplate reader (SpectraMax Gemini EM, Molecular Devices, USA).
Determination of Interleukin-8 (IL-8) Secretion
To quantify tumor necrosis factor-α (TNF-α)-induced IL-8 secretion in HT-29 cells, 1 × 106 cells/well were seeded in a 6-well plate and cultured for 24 h. The cells were treated with 200 µg/mL KPB in the presence of 10 ng/mL TNF-α (PeproTech, USA) and incubated for an additional 24 h. The culture medium was collected and centrifuged at 5,000 rpm for 10 min; the supernatant was harvested for analysis. IL-8 secretion was quantified using an IL-8 ELISA kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions.
Experimental Animals
Six-week-old male C57BL/6N mice were obtained from Daehan BioLink Co., Ltd. (Eumseong, Korea) and acclimated for one week in an animal facility maintained at 23 ± 1 °C and 50 ± 5% relative humidity under a 12-h light/dark cycle with ad libitum access to standard laboratory chow (2918C Diets, Envigo, USA) and water. The mice were randomly assigned based on body weight into four groups (n = 8/group): the normal control (NC), DSS group (2.5% DSS; DSS), low-dose KPB (2.5% DSS + KPB 10 mg/kg; KPB-L), and high-dose KPB (2.5% DSS + KPB 100 mg/kg; KPB-H) groups.
Colitis was induced by providing 2.5% DSS (MW 36-50 kDa, MP Biomedicals) dissolved in sterile drinking water ad libitum for five days, followed by sterile drinking water for the remainder of the experiment. KPB suspended in PBS was orally administered once daily for two weeks. The NC and DSS groups received 200 µL of PBS as the vehicle control.
All animal experiments adhered to relevant regulations governing the care and use of laboratory animals and were approved by the Institutional Animal Care and Use Committee (IACUC) of the Hongcheon Institute of Medicinal Herb (Approval No. HIMH A23-03).
Assessment of Disease Activity Index (DAI)
Mice were weighed at the same time each morning, and mice were placed in clean individual cages for fecal sample collection. The presence of fecal occult blood and the severity of diarrhea were assessed using Coloscreen III test kit (Helena Laboratories, USA). The DAI score was calculated based on body weight loss, stool consistency, and fecal occult blood, according to the score criteria presented in Table 2 (Berberat et al., 2005).
Table 2
Criteria for disease activity index (DAI) in DSS-induced colitis
| Score | Body weight loss | Stool consistency | Occult/gross bleeding |
| 0 | None | Normal | Normal |
| 1 | 0-10% | Watery | - |
| 2 | 10-15% | Loose stools | Hemoccult + |
| 3 | 15-20% | - | |
| 4 | > 20% | Diarrhea | Gross bleeding |
Histological Analysis of Colon Tissue
After the 14-day treatment period, animals were deeply anesthetized by isoflurane inhalation, and blood was collected via the orbital sinus. The animals were then euthanized by exsanguination, followed by organ collection. The colon extending from the cecum to the rectum was resected, and its length was measured. The colon was washed with PBS (pH 7.4), weighed, and fixed in 10% formalin using the Swiss-roll method. Tissues were paraffin-embedded, sectioned at 5 µm thickness, and stained with hematoxylin and eosin (H&E). Histopathological scoring of colonic tissue was performed according to the criteria presented in Table 3, evaluating inflammatory cell infiltration, epithelial injury, and mucosal ulceration (Erben et al., 2014).
Table 3
Criteria for scoring DSS-induced colonic inflammation
Measurement of IL-6 and IL-22 Levels
Plasma was separated from the collected blood samples by centrifugation at 4,000 rpm for 15 min. IL-6 and IL-22 levels were quantified using ELISA kits (R&D Systems, USA) according to the manufacturer’s instructions.
Statistical Analysis
All results are presented as the mean ± standard deviation (SD). Statistical analysis was conducted using GraphPad Prism version 7.05 (GraphPad Software, San Diego, CA, USA). Data collected at single time points were analyzed by one-way analysis of variance (ANOVA) as the independent variables.
Changes in body weight, diarrhea index, fecal occult blood index, and DAI during the treatment period were analyzed using two-way repeated-measures ANOVA with treatment group and time as factors. Dunnett's multiple comparison test was used for post hoc comparisons, and statistical significance was set at p < 0.05.
Results and Discussion
Effect of KPB on Cell Viability
The cytotoxicity of KPB in HT-29 cells was assessed using a WST assay. KPB showed no cytotoxic effects at concentrations up to 800 µg/mL (Fig. 2A). However, treatment with 1600 µg/mL significantly reduced cell viability to 68.12 ± 7.26%. Therefore, 200 µg/mL was selected as the treatment concentration for subsequent in vitro experiments.
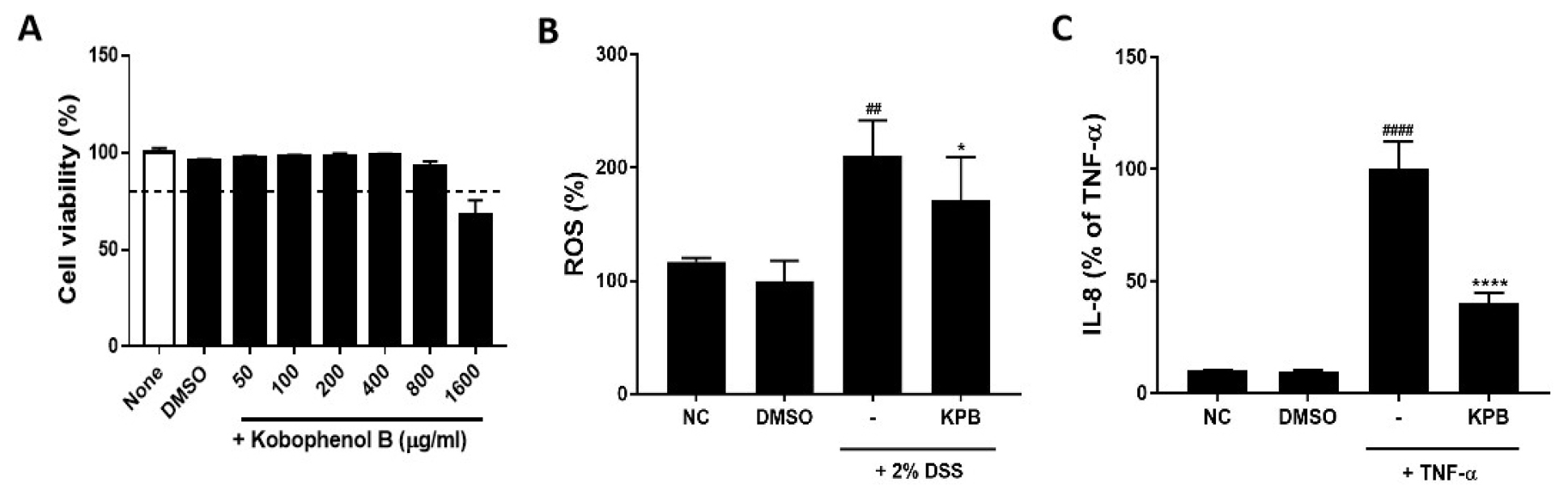
Fig. 2.
Effects of Kobophenol B on cell viability, reactive oxygen species (ROS) production, and Interleukin-8 (IL-8) secretion in HT-29 cells. (A) Cell viability assessed by WST assay; (B) intracellular ROS production in 2% DSS-treated HT-29 cells; (C) IL-8 secretion induced by TNF-α (10 ng/mL). Data are expressed as mean ± SD (n = 3). Statistical significance was analyzed using one-way ANOVA followed by Dunnett’s multiple comparison test. ##p < 0.01, ####p < 0.0001 vs. the normal control (NC) group; *p < 0.05 vs. the DSS group; ****p < 0.0001 vs. the TNF-α group.
Inhibitory Effect of KPB on ROS Production
To assess intracellular oxidative stress, HT-29 cells were treated with 2% DSS, and ROS production was measured using DCFH-DA (Fig. 2B). ROS production in the 2% DSS-treated group increased significantly to 210.7 ± 30.9% compared with control cells (p < 0.01). However, co-treatment with KPB (200 µg/mL) significantly decreased DSS-induced ROS production to 171.5 ± 37.7%, corresponding to an approximate 18.6% reduction compared with DSS-treated cells (p < 0.05). These findings indicate that KPB suppresses DSS-induced oxidative stress in HT-29 cells.
As a stilbene oligomer and resveratrol derivative, KPB has been reported to exhibit potent free-radical-scavenging activity, which is attributed to its conjugated structure and multiple phenolic hydroxyl groups (Duta-Bratu et al., 2023). ROS serve as key regulators of the NF-κB and MAPK signaling pathway. Therefore, the antioxidant capacity of KPB may contribute to its anti-inflammatory effects by modulating ROS-mediated inflammatory responses (Liu et al., 2025).
Inhibitory Effect of KPB on IL-8 Production
Treatment of HT-29 colonic epithelial cells with TNF-α (10 ng/mL) induced an inflammatory response, including a statistically significant increase in IL-8 secretion compared with the NC group (Fig. 2C). TNF-α rapidly activates the NF-κB signaling pathway in intestinal epithelial cells, thereby enhancing the secretion of neutrophil-inducing chemokines such as IL-8 (Bai et al., 2004). IL-8 promotes neutrophil infiltration into the intestinal mucosa, amplifying the local inflammatory response and contributing to disruption of the intestinal epithelial barrier—a critical factor in IBD pathogenesis (Ko and Auyeung, 2014).
Conversely, treatment with KPB (200 µg/mL) markedly reduced the TNF-α-induced increase in IL-8 secretion to 40.08 ± 4.89% (p < 0.0001). These findings indicate that KPB effectively suppresses TNF-α-induced IL-8 production in HT-29 cells. Previous studies have demonstrated that KPB exerts anti-inflammatory activity by inhibiting the NF-κB and MAPK signaling pathways (Cho et al., 2018). The observed inhibitory effect on IL-8 production is consistent with these reported anti-inflammatory mechanisms. Therefore, KPB may serve as a promising natural compound for the prevention or amelioration of chronic inflammatory diseases such as IBD by modulating the excessive secretion of pro-inflammatory cytokines in the intestinal mucosa.
Effect of KPB on Disease Activity Index (DAI)
Following administration of 2.5% DSS for 5 days, mice began to exhibit body weight loss from Day 6 (Fig. 3B). In DSS-induced colitis models, body weight loss is a typical clinical indicator reflecting malabsorption caused by intestinal epithelial damage and systemic inflammation (Cooper et al., 1993). On Day 9, the DSS group exhibited the largest reduction in body weight (20.67%), followed by the KPB-L (18.44%) and KPB-H (14.54%) groups. Notably, the KPB-H group showed a significant attenuation of body weight loss compared with the DSS group between Days 9 and 12 (p < 0.05). Although the KPB-L group also showed a tendency toward reduced body weight loss, the difference was not statistically significant.
The stool consistency score increased sharply on Day 6 in the DSS group (p < 0.0001) and reached a peak on Days 7 and 8. During this peak period, the KPB-treated groups showed significantly lower stool consistency scores compared with the DSS group (Fig. 3C). Occult blood appeared as early as Day 1, and the fecal occult blood score increased markedly from Day 3 of DSS treatment, reaching a peak on Day 5 (Fig. 3D, p < 0.001). The mean fecal occult blood scores were 3.60 ± 0.85 for the DSS group, 3.50 ± 0.09 for the KPB-L, and 3.00 ± 1.07 for the KPB-H. Although the KPB-treated groups showed a tendency toward lower fecal occult blood scores, the differences were not statistically significant. Diarrhea and bloody stools induced by DSS are clinical indicators of intestinal mucosal injury and impaired intestinal barrier function. The reduced severity of these symptoms in the KPB-treated groups suggests that KPB may attenuate DSS-induced intestinal mucosal damage.
Based on body weight loss, stool consistency, and fecal bleeding scores, the DAI score in the DSS group peaked at 7.70 ± 2.21 on Day 8 (p < 0.01). In contrast, the DAI scores were reduced to 6.13 ± 1.96 in the KPB-H and 5.50 ± 1.93 in the KPB-L group (Fig. 3E). In particular, the KPB-H group showed a significant reduction in DAI score compared with the DSS group (p < 0.01), indicating that KPB alleviated the severity of DSS-induced acute colitis.
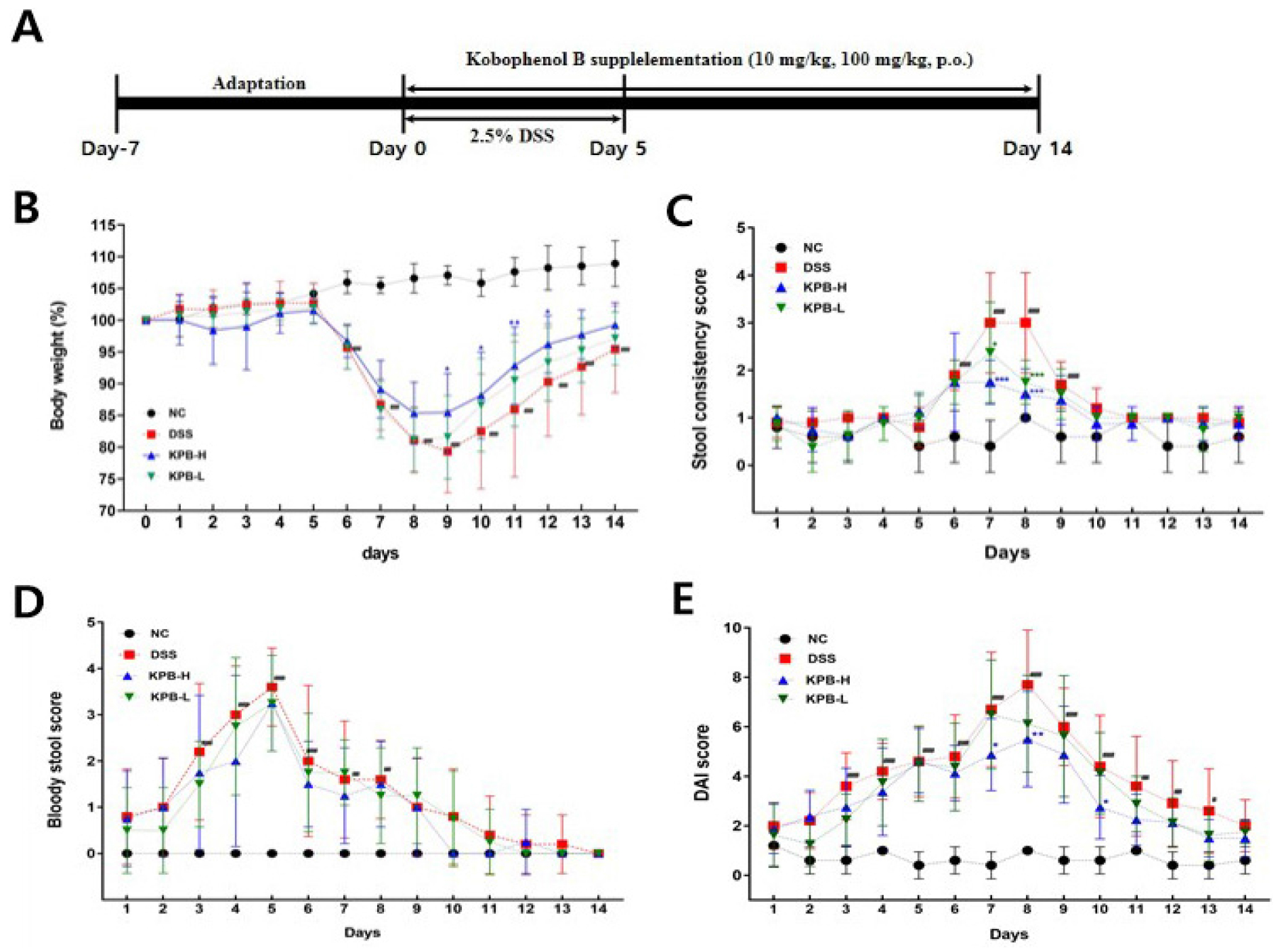
Fig. 3.
Effects of Kobophenol B on clinical parameters in DSS-induced acute colitis mice. Acute colitis was induced by administration of 2.5% DSS in drinking water for 5 days, and Kobophenol B was orally administered for 14 days throughout the experimental period. (A) Experimental design, (B) body weight change (%), (C) stool consistency score, (D) bloody stool score. Data are expressed as mean ± SD (n = 8). Statistical significance was analyzed using two-way repeated measures ANOVA followed by Dunnett’s multiple comparison test. #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001 vs. the normal control (NC) group; **p < 0.01, ***p < 0.001 vs. the DSS group.
Effects of KPB on DSS Intake, Spleen Weight, Colon Weight, and Colon Length
The average daily DSS intake was 4.32 ± 1.09 mL/day in the NC group and 4.69 ± 1.18 mL/day in the DSS group, indicating an 8.6% increase in DSS solution intake in the DSS group; however, this difference was not statistically significant (Fig. 4A). Therefore, these findings suggest that differences in DSS intake did not influence the severity of colitis.
In the DSS-induced colitis model, spleen weight increased significantly in the DSS group (110.99 ± 28.47 mg), representing a 69.2% increase compared with the NC group (65.60 ± 5.74 mg, p < 0.01; Fig. 4B). Splenomegaly in this model reflects systemic inflammatory responses and immune cell activation (Yan et al., 2009). Notably, KPB treatment reduced spleen weight relative to the DSS group, with decreases of 10.58% in the KPB-H group (99.25 ± 26.73 mg) and 6.32% in the KPB-L group (103.98 ± 17.35 mg), suggesting that KPB may alleviate local inflammation and systemic immune responses.
Colon length, a representative pathological indicator of DSS-induced colitis, typically decreases with increasing inflammation and intestinal fibrosis (Perše and Cerar, 2012). In this study, colon length was 6.57 ± 0.42 cm in the NC group and 5.55 ± 0.35 cm in the DSS group, indicating a significant reduction of 15.6% (Fig. 4D, p < 0.001). The KPB-treated groups exhibited colon lengths of 5.74 ± 0.43 cm (KPB-H) and 5.58 ± 0.27 cm (KPB-L); however, these changes were not statistically significant compared with the DSS group.
The colon weight per unit length increased substantially in the DSS group (51.16 ± 5.38 mg/cm), representing a 187.7% increase relative to the NC group (27.25 ± 1.66 mg/cm, p < 0.0001; Fig. 4E). In the DSS-induced colitis model, this parameter reflects mucosal edema and inflammatory cell infiltration (Murthy et al., 1993). The KPB-H group exhibited a significant 15.28% reduction (43.34 ± 6.13 mg/cm) compared with the DSS group (p < 0.01), suggesting that KPB may attenuate mucosal edema and inflammatory cell infiltration in colonic tissue.
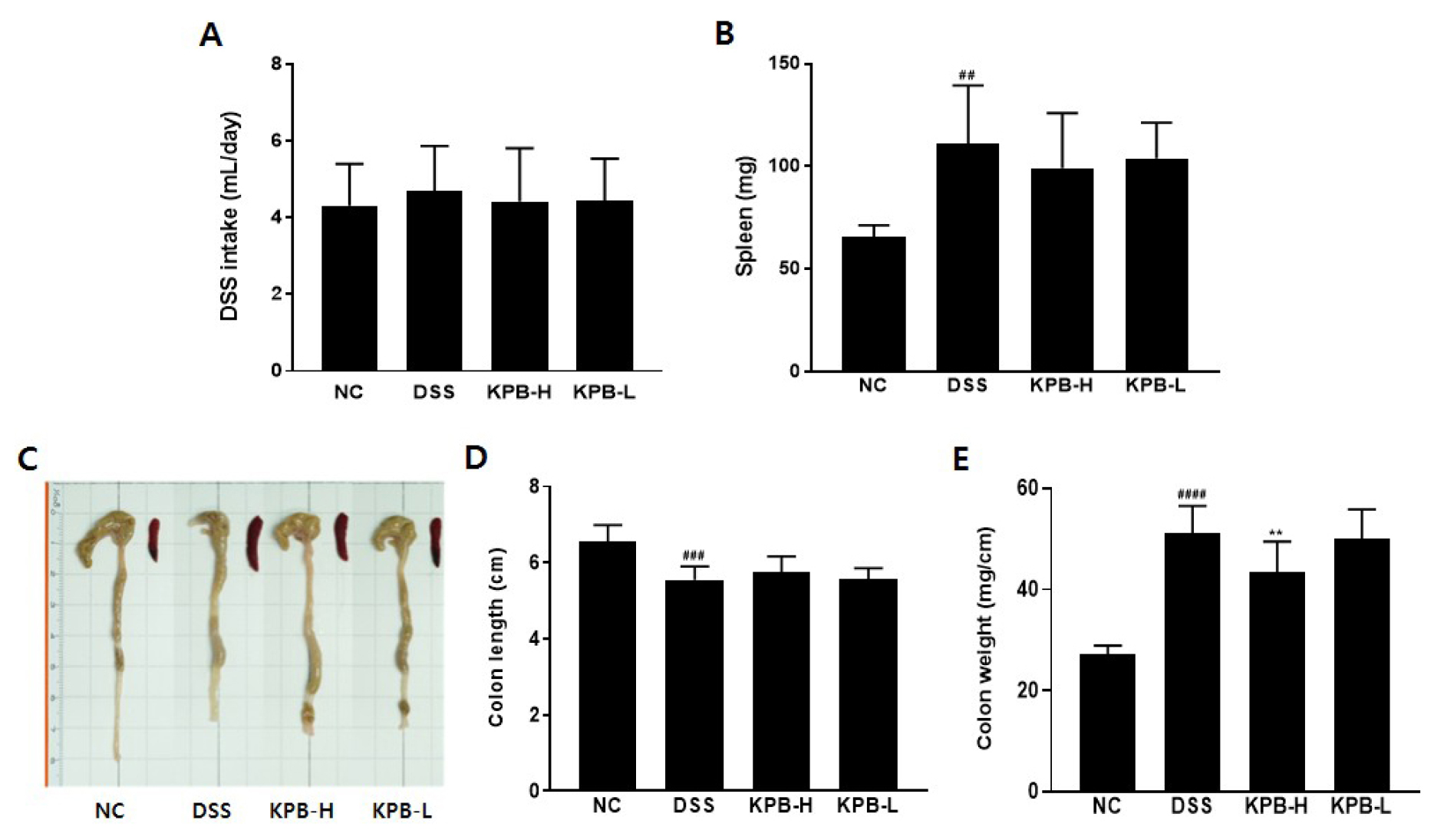
Fig. 4.
Effects of Kobophenol B on spleen weight and colon morphology in DSS-induced colitis mice. Acute colitis was induced by administration of 2.5% DSS in drinking water for 5 days, and Kobophenol B was orally administered for 14 days. (A) Representative images of the colon and spleen, (B) DSS intake, (C) spleen weight, (D) colon length, and (E) colon weight per unit length. Data are expressed as mean ± SD (n = 8). Statistical significance was analyzed using one-way ANOVA followed by Dunnett’s multiple comparison test. ##p < 0.01, ###p < 0.001, ####p < 0.0001 vs. the normal control (NC) group; **p < 0.01 vs. the DSS group.
Histopathological Analysis of Colon Tissue
H&E staining was performed to evaluate colonic mucosal damage and inflammatory cell infiltration in a DSS-induced colitis model (Fig. 5). In the DSS-treated group, localized or extensive ulcerative lesions were observed in the mucosal layer, showing histopathological features typical of acute colitis. Inflammatory cells infiltrated beyond the mucosal layer into the submucosa and muscularis propria (Fig. 5D). These findings are consistent with previous studies indicating that DSS directly disrupts the intestinal epithelial barrier, triggering an acute inflammatory response results in inflammatory cell infiltration and disruption of tissue architecture (Wirtz et al., 2007).
The KPB-treated group exhibited inflammatory cell infiltration and ulcerative mucosal lesions similar to those observed in the DSS group, with no significant improvement in histological scores (Fig. 5A-C). In the present study, the lack of a significant reduction in histological scores following KPB treatment may be attributed to the limited ability to structurally restore the rapidly disrupted intestinal barrier induced by DSS within a short experimental period. This finding is consistent with previous studies reporting that polyphenolic compounds effectively suppress inflammatory mediators but that a longer recovery period is required for the complete restoration of epithelial tissue (Larrosa et al., 2009). Thus, KPB is considered to primarily regulate acute immune responses and modulate clinical severity rather than directly promote rapid epithelial regeneration in the acute DSS-induced environment. Further studies using recovery-phase or long-term treatment models are warranted to evaluate its potential tissue-regenerative effects.
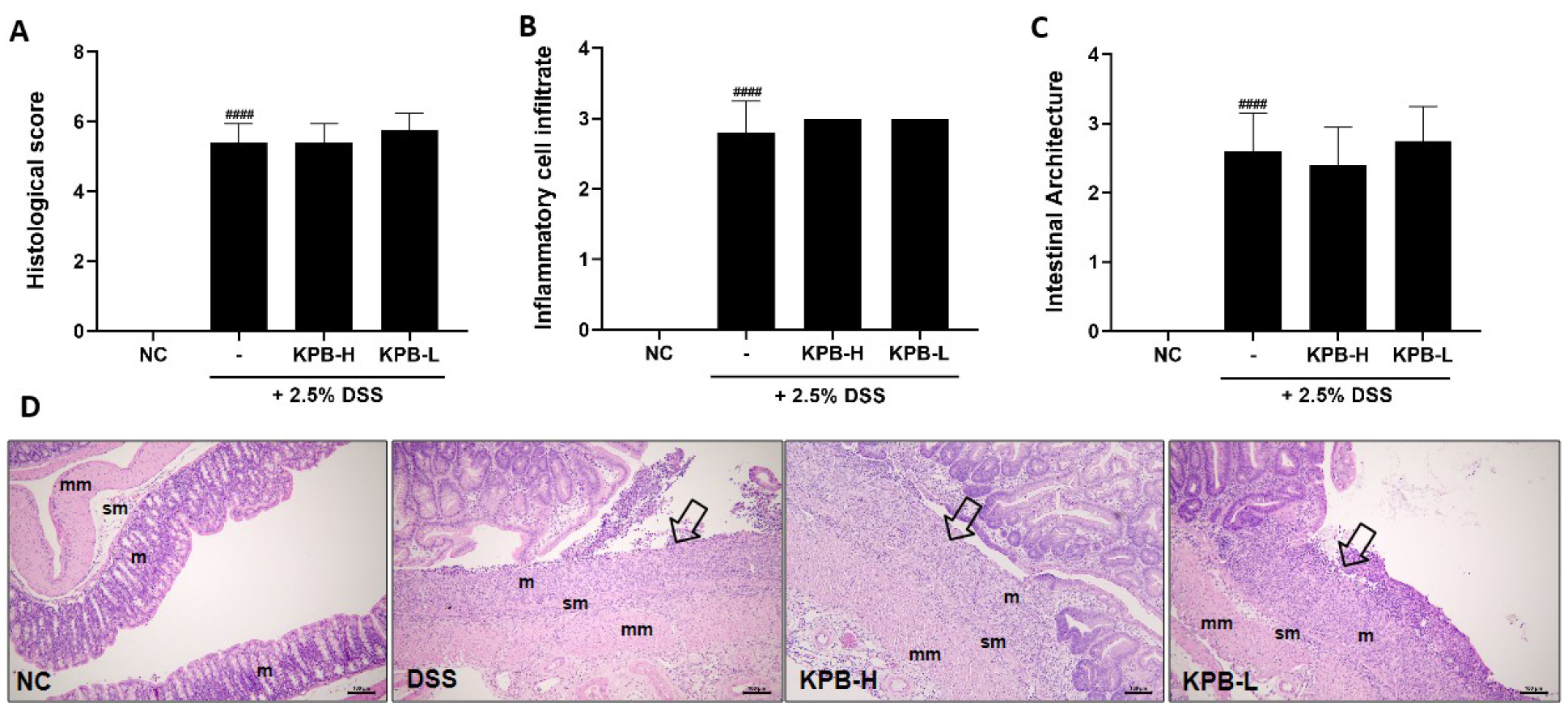
Fig. 5.
Effects of Kobophenol B on histopathological changes in the colon of DSS-induced colitis mice. (A) Histological score, (B) inflammatory cell infiltration score, (C) intestinal architecture score, and (D) representative hematoxylin and eosin (H&E)-stained colon sections (scale bar = 100 μm). m, mucosa; sm, submucosa; mm, muscular layer. Arrows indicate ulcerative lesions. Data are expressed as mean ± SD (n = 8). Statistical significance was analyzed using one-way ANOVA followed by Dunnett’s multiple comparison test. ###p < 0.0001 vs. the normal control (NC) group.
Effects of KPB on IL-6 and IL-22 levels
Administration of 2.5% DSS significantly increased serum IL-6 concentrations in the DSS group (100.00 ± 52.88%) compared with the NC group (2.83 ± 2.75%), representing an approximately 35-fold increase, confirming that DSS induced a systemic inflammatory response (Fig. 6A). In contrast, KPB treatment attenuated the DSS-induced increase in serum IL-6 levels, with values of 52.87 ± 4.63% in the KPB-H group and 54.95 ± 38.40% in the KPB-L group. Notably, the KPB-H group showed a significant reduction compared with the DSS group (p < 0.05).
IL-22, a cytokine primarily secreted by Th17 cells and innate lymphoid cells, promotes regeneration of damaged intestinal mucosa and epithelial cell proliferation. Increased IL-22 expression is associated with the severity of intestinal mucosal injury (Zindl et al., 2013). In this study, serum IL-22 levels in the DSS group (100.00 ± 37.95%) were more than fourfold higher than those in the NC group (21.86 ± 17.30%; p < 0.001), indicating that DSS simultaneously activated mucosal damage and regenerative responses (Fig. 6B). The KPB-H and KPB-L groups showed concentration-dependent suppression of IL-22 levels to 53.29 ± 25.84% and 66.61 ± 28.10%, respectively (p < 0.01). These findings suggest that KPB may attenuate the inflammatory response and subsequently reduce excessive regenerative signaling.

Fig. 6.
Effects of Kobophenol B on inflammatory cytokine levels in the plasma of DSS-induced colitis mice. Plasma concentrations of (A) IL-6 and (B) IL-22 were measured by ELISAs. Data are expressed as mean ± SD (n = 8). Statistical significance was analyzed using one-way ANOVA followed by Dunnett’s multiple comparison test. ###p < 0.0001 vs. the normal control (NC)) group; *p < 0.05, **p < 0.01 vs. the DSS group.
Summary
This study evaluated the therapeutic effects of KPB, isolated and purified from C. pumila Thunb., on inflammatory bowel disease (IBD). In vitro experiments demonstrated that KPB significantly reduced ROS production and the secretion of the inflammatory chemokine IL-8 induced by DSS or TNF-α in HT-29 cells. In vivo, oral administration of KPB (10 and 100 mg/kg) for 14 days in a 2.5% DSS-induced acute colitis mouse model significantly improved DAI, including body weight loss, stool consistency and bloody stools, while alleviating splenomegaly and reducing colon weight per unit length. Furthermore, KPB treatment suppressed the DSS-induced increase in IL-6 and IL-22 levels in a concentration-dependent manner.
Collectively, these results suggest that KPB exerts significant anti-inflammatory effects in the DSS-induced colitis model and may serve as a potential natural product-derived therapeutic candidate for IBD.
