

Introduction
Sesame
11S Globulin (Legumin)
Almond
Amandin, 11S Globulin from Almond
Pumpkin
Cucurbitin, 11S Globulin from Pumpkin Seeds
Bioactive Peptides
Psi Blast Based Homology
In-silico analysis for identification of proteins with bioactive peptides
Alpha Amylase and Alpha Glucosidase Inhibiting Peptides
Antihypertensive Peptides
Globulin protein of sesame seeds, amandin protein of almond and cucurbitin protein of pumpkin seeds
Angiotensin-converting Enzyme (ACE)
Vasodilating Effect of Nitric Oxide
Mechanism of NO
Spermine NONOATE and NO Donor
Conclusion
Future Perspectives
Introduction
Plant proteins are important functional ingredients in many processed food products. Particularly globular proteins from various sources play an important role in many food products, due their contribution to food texture (McClements and Gumus, 2016). The seed storage proteins have been traditionally classified through the sequential extraction of crushed and defatted seeds by a series of aqueous and non-aqueous solvents (Marcone et al., 2002). The fraction extracted with water is defined as albumins, the fraction with dilute salt as globulins, the fraction with ethanol as prolamines, and the fraction with acid or alkali as glutelins. In cereal grains the main storage proteins are usually the alcohol soluble prolamines, whereas, in non-cereal grains the more nutritiously balanced salt soluble globulins predominate (Ali, 2015; Fageer, 2015; Janssen et al., 2017). Globulins can be subdivided into two distinct classes termed 7S and 11S on the basis of their sedimentation coefficient (Czubinskiet al., 2015). Dicotyledonous plants have been found to favors the presence of the 11S globulin form (Sharma et al., 2017; Vajravijayan et al., 2017). 11S globulins are found to occur in the 300 kDa range whereas 7S globulins are generally less abundant and found to occur in the 180 kDa range.
The globulins storage proteins are classified into two broad groups, on the basis of their sedimentation coefficients: 7S Vicilin-type and 11S Legumin-type. The majority of the storage globulin proteins are soluble in dilute salt solution but insoluble in water. The two main reserve proteins of soybean that are used in the food industry are 7S β-conglycinin and 11S glycinin globulins. The 7S globulin is a trimeric glycoprotein (141-170 KDa) composed of three subunits, α (57 kDa), α’ (58 kDa) and β (42 kDa), associated by hydrophobic interactions (Tan-Wilson and Wilson, 2012; Marambe et al., 2012; Bhushan and Dixit, 2012; Jyothi, 2007). Sesame (Sesamum indicum) is the most ancient oil seed known and used by humans as a food source and it is mainly cultivated for its oil, whole or dehulled seeds are used in confectionery foods (Sumaya Hassan Abdel Rahim, 2015). It is also known as gingely, beniseed, sim sim and til. This annual seed crop has been cultivated for centuries, in the developing countries of Asia and Africa, for its high quality oil and protein (Sediqi, 2012). Sesame oil is different from all other vegetable oils in many chemical, biological and physiological properties. These properties are due to the presence of endogenous unsaponifiable constituents viz, sesamol, sesamin, and sesamolin. Sesamin is reported to possess in vivo hypocholesterolemic activity and suppressive activity against chemically induced cancer (Kochhar, 2011).
Almond (Prunus dulcis), belongs to the Rosaceae family that also includes apples, pears, prunes, and raspberries. Almond is one of the most popular tree nuts on a worldwide basis and ranks number one in tree nut production (Esfahlan et al., 2010). They are typically used as snack foods and as ingredients in a variety of processed foods, especially in bakery and confectionery products (Monteiro et al., 2013). Extracts of whole almond seed, brown skin, shell, and green shell cover (hull) possess potent free radical-scavenging capacities (Sang et al., 2002). In addition, almonds, when used as snacks and in diets of hyperlipidemic subjects, significantly reduced coronary heart disease (Jamshed et al., 2015) A long-term supplementation of almond showed spontaneous nutrient modification of an individual’s habitual diet that closely matched the recommendations to prevent cardiovascular and other chronic diseases (Abazarfard et al., 2014; Chen et al., 2015).
Pumpkin (Cucurbita maxima) belongs to the family cucurbitaceae. It is a leafy green vegetable (Kindinew, 2015; Olasantan, 2007). Fruits are variable in size, color, shape, and weight. Pumpkin has received considerable attention in recent years because of the nutritional and health protective value of the proteins from the seeds. Pumpkin seed proteins, beside their wide use as food ingredients, have pharmacological activities too such as antidiabetic (Chonoko and Rufai, 2011; Singh, 2012; Tomar et al., 2014), antifungal (Abdel-Rahim et al., 2015; Libo et al., 2014; Qian, 2013), antibacterial and anti-inflammation activities (Al-Okbi et al., 2017; Perez Gutierrez et al., 2016). Pumpkin storage proteins are 2S albumins and 11S globulins (cucurbitin) localized in the protein bodies (Gallardo et al., 2016; Hegedus et al., 2015; Shimada et al., 2017). Alpha amylase inhibitors have potential roles in controlling blood sugar levels. Alpha glucosidase inhibitors are used as oral anti diabetic drugs for treating type 2 diabetes mellitus. They act by preventing the digestion of carbohydrates such as starch. Carbohydrates are normally converted into simple sugars which can be absorbed through the intestine (Nair et al., 2013). Alpha glucosidase inhibitors act as competitive inhibitors of alpha glucosidase enzyme which is needed to digest carbohydrates. The intestinal alpha glucosidases hydrolyze complex carbohydrates to glucose and other monosaccharides in the small intestine. Inhibition of these enzyme systems helps to reduce the rate of digestion of carbohydrates. Less amounts of glucose is absorbed because the carbohydrates are not broken down into glucose molecules (Panwar et al., 2014).
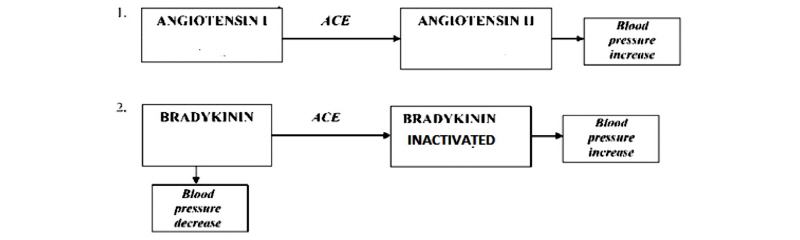
Hypertension is a growing undesired symptom that damages health and threatens mostly the developed societies. It is one among the major independent risk factors for atherosclerosis, stroke, myocardial infarction, and end stage renal disease. Angiotensin I-converting enzyme, adipeptidyl-dipeptidase, is the key enzyme that functions in the rennin angiotensin system to increases blood pressure (Hermida et al., 2016; Olsen et al., 2016). ACE increases blood pressure by both converting the inactive decapeptide Angiotensin I to the potent vasoconstrictor Angiotensin II and inactivating the vasodilator bradykinin. Inhibition of ACE is considered the first line of therapy for hypertension and atherosclerosis. ACE inhibitors have short peptides that bind tightly to the active site of ACE competing with Angiotensin I for occupancy (Braunwald, 2015).
The synthetic inhibitors of ACE have established in the therapy of hypertension and congestive heart failure. Synthetic ACE inhibitors are remarkably effective, but they cause adverse side effects, and therefore, the trend has been toward developing safer, natural ACE inhibitors. ACE inhibitor peptides derived from daily dietary food proteins would be useful in the development of a novel functional food additive and present a healthier and natural alternative to ACE inhibitor drugs (Te Riet et al., 2015).
Seed cake, by product of oil industry is used as cattle feed because of high content of protein. Sesame, almond and pumpkin seed cake are rich in protein. In this study globulin protein was extracted from the defatted seeds. Alpha amylase inhibiting and antihypertensive peptides were identified in 11S globulin protein of sesame, almond and pumpkin seeds by in silico analysis. Alpha amylase inhibiting activity and antihypertensive activity of 11S globulin protein was evaluated (Mäkinen, 2014; Pojić et al., 2014).
The current review focus on globulin, amandin, cucurbitin protein from sesame seeds, sweet almond seeds and pumpkin seeds. Protein by column chromatography; further to characterize 11S globulin, amandin and cucurbitin protein by SDSPage and in addition compare the alpha amylase inhibiting activity and anti-hypertensiveactivity of 11S globulin, amandin and cucurbitin whole protein.
Sesame
Sesame crop has been grown for centuries owing to its high content of excellent quality oil and protein (Gharby et al., 2017; Pathak et al., 2014). The amino acid composition of the sesame seeds is unique and unusual among the oilseed proteins, due to its high content of sulphur- containing amino acids (methionine and cysteine) and low content of lysine (Khan et al., 2016; Laohakunjit et al., 2017; Teh et al., 2017). It is one of the earliest condiment and crop grown for edible oil. It is consumed directly as sweetmeat and snack. The seed has been called “Queen of the oil seed crops” due to its high oil yield and its quality (Miraj and Kiani, 2016). The dehulled and defatted meal can be used in food products as a protein and tryptophan/methionine supplement (Gandhi et al., 2007; Widyarani, 2016). Sesame seed contains 40-60% oil, 20-25% protein, 20-25% carbohydrate and 5-6% ash (Couch et al., 2017). Extraction of oil has led to increased protein content of defatted sesame meal (41.15-49.58%) (Asghar and Majeed, 2013). Although the seed contains anti nutritional factors like phytates and oxalates, they are significantly minimized during processing. Defatted sesame meal contains more sugar and is generally utilized as animal feed and often as manure (Suliman, 2015). This meal has a great potential in combating the protein calories malnutrition because of its high quality and quantity of protein. However there is a need to process the meal carefully for human consumption (Chatterjee, 2016). The functional properties of sesame protein concentrate. Functional properties of protein are important in food processing and food formulation. Some of the functional properties are solubility, water and oil holding capacity, foaming capacity and stability, gelation, bulk density and viscosity (Elleuch et al., 2014; Gómez-Arellano et al., 2017). Sesame protein is used for the preparation of sweets, confectionary and bakery products (Laohakunjit et al., 2017). It is used as nutritional supplements, functional ingredients, and flavor enhancers in foods. Protein can be used as additives, as food texture enhancers. When comparing emulsifying properties and protein solubility of a sesame protein isolate with those of a soybean protein isolate, it was found the emulsifying activity index (EAI) of the sesame protein isolate were better than those of the soybean protein isolate at pH 4 and 7. For protein solubility, the sesame protein isolate was about 15 times more soluble than the soybean protein isolate at acidic pH 2-4, but not in neutral and alkaline condition (Hijar, 2013; Soltero, 2013).
11S Globulin (Legumin)
The 11S globulins or legumin-like proteins are members of the cupin superfamily which are characterized by a 6 stranded β–barrel conformation (Singh et al., 2015; Pereira, 2013). They are typically hexameric -360 kDa proteins in which each monomeric subunit is comprised of an acidic 40–42 kDa polypeptide that is disulfide-linked to a 20 kDa basic polypeptide (Albillos et al., 2008; Sharma et al., 2010). In the plant, seed storage proteins serve as amino acid reserves for the developing seedling and act as a carbon and nitrogen source during germination (Aragão et al., 2015; Ben Mrid et al., 2017; Yoneyama et al., 2016). Seed storage proteins are abundant in the seed and it is estimated that they can account for -40% of the total protein (Foley et al., 2015; Riascos et al., 2016).
Almond
Almond belong to the genus Prunus and the subgenus Amygdalus, within the Rosaceae family. Almond tree are broadly be divided into two types, bitter and sweet (Eisenman, 2015; Woldring, 2015). The cultivated sweet almond (Prunus dulcis) is a nutritionally important and valuable crop grown in many temperate and sub-tropical regions in the world, both for domestic consumption and for trade (Gradziel, 2017; Parrotta et al., 2015). Sweet almonds have been cultivated for thousands of years and valued for both food and medicinal uses. The oil content of dried sweet almond kernel is 50-60% (Yanmife, 2016). The cake of sweet almond remaining after oil extraction contains 39-47% protein and 10-18% oil (Khan and Abourashed, 2011). Bitter almonds has oil with lower yield 40-45% and sometimes as low as 20% (Akubude and Nwaigwe, 2016). Bitter almonds are used primarily in the production of flavor extracts (Akpakpan and Akpabio, 2012). Almonds are nutrient dense foods that can be a valuable plant source of lipids and protein in the diet (Inarejos-García et al., 2010). The contents of dietary fiber, vitamin E, phytosterols and several key micronutrients found in almonds contribute to a healthy nutrient profile. Almond skins as a rich source of bioactive polyphenols is further enhancing the health profile of this nut (Alasalvar and Bolling, 2015; Brown et al., 2016; Prgomet et al., 2017). The Prunus genus is reported to have interesting biological properties such as sedative, anti-inflammatory, antihyperlipidemic, anti-tumoural and antioxidant activities (Esfahlan et al., 2010).
Amandin, 11S Globulin from Almond
Amandin is an 11S globulin legumin-like protein that is hexameric in form and composed of individual polypeptides (Bojórquez-Velázquez et al, 2016; de la Cruz et al., 2017). Each monomeric subunit, like those of other 11S globulins, is comprised of a large 40-42 kDa acidic chain and a small 20-22 kDa basic chain that are linked by a disulfide bond (McNulty, 2006; Wanasundara and McIntosh, 2013).Two isoforms have been identified in almond, prunin 1 and prunin 2, which comprise native prunin (Niezgoda, 2015). Analyses of almond protein extract has been extensively carried out and it was found that prunin is a major component of the nut, accounting for up to 65% of the protein in the soluble extract (da Costa, 2013). Analysis revealed that the overall structure of amandin is typical of the 11S globulin class of seed storage proteins, as it is composed of two prunin trimers that associate to form the native hexameric structure (Chhabra, 2013).
Pumpkin
Pumpkin seeds, commonly known as ‘pepitas’, are flat, encased in yellow-white husk (Amin and Thakur, 2013). Pumpkin seeds contain phytocompounds which have significant anti-nutritional effects (Gorrepati et al., 2015). These compounds include oxalate, phytate, nitrate, cyanide, etc. Pumpkin seeds are consumed directly as snack food (Babita, 2013). It was reported by (Longato et al., 2017) that the nutritional value of seeds increases after roasting. Roasting basically increases the sterols andk vitamin E content. Pumpkin seeds are rich in protein content (25-52%) (Hassan et al., 2016). It was reported by (Bhoi 2014) that oil content is also high, ranging from 40-60%. Out of this, up to 60.8% is contributed from fatty acids oleic acid (up to 46.9%), linolenic acid (up to 40.5%), palmitic and stearic acid up to 17.4%, the ratio of monounsaturated to polyunsaturated acids 0.60 to 0.75 g. Approximately 1% each of phytosterols, squalene and chlorophyll pigment is present. Phytosterols are present in free and bound form. Minerals (including selenium, zinc, calcium, copper, iron, manganese, phosphorus and potassium) are present at 4-5%. Pectin content is 30% (Amin and Thakur, 2013). Pumpkin seeds are consumed directly for human consumption as a snack food in many cultures throughout the world (Abdel-Rahman, 2006). The kernels of pumpkin seeds have been utilized as additives to some food dishes (Amin and Thakur, 2013). Seeds belonging to Cucurbitaceae family are known to be as rich in oil as soybean, cottonseed, or corn (Gorrepati et al., 2015). Pumpkin seed extract has been reported to have antidiabetic, antitumor, antibacterial, antihypertensive, antifungal, antiinflammatory, anticancer, antimutagenic, and antioxidant activities. It has also been found to have bone protection, serum cholesterol-lowering and relief from anxiety effects (Keservani et al., 2016). Defatted pumpkin seed cake, which is a by-product of oil extraction, is usually used only as animal feed (Gorrepati et al., 2015). However, pumpkin oil cake is rich in protein which could be used for human nutrition. The protein exploitation from pumpkin oil cake has been described as a way to increase the value of this agricultural by-product (Keservani et al., 2016). Oilseed proteins are used in food formulations as nutrition supplements, as well as functional agents (Gorrepati et al., 2015). Protein bodies are widely distributed in the seeds of higher plants. They are membrane -bound organelles in which various seed proteins are stored. Three major proteins, 11S globulins, 7S globulins and 2S albumin, are present in the protein bodies of pumpkin seeds. 11S globulin (cucurbitin) forms crystalloids in the protein bodies, while 7S globulin and 2S albumin are located in the matrix of the protein bodies (Zhao et al., 2016).
Cucurbitin, 11S Globulin from Pumpkin Seeds
Cucurbitin, the major storage protein in pumpkin seed belongs to 11S globulin family, was described many years ago by Osborne (Narasimhan, 2014). It is a hexamer with a molecular size of about 350 kDa (Seiter, 2009). Hexamer protein contains six monomer subunits. Each monomeric subunit is comprised of an acidic polypeptide (33 kDa) and a basic polypeptide (22 kDa).
Bioactive Peptides
Bioactive peptides have been defined as peptides with hormone- or drug like activity that eventually modulate physiological function through binding interactions to specific receptors on target cells leading to induction of physiological responses. According to their functional properties, bioactive peptides may be classified as antimicrobial, antithrombotic, antihypertensive, opioid, immunomodulatory, mineral binding and anti-oxidative. These peptides play an important role in human health.
Psi Blast Based Homology
BLAST program, searches for sequences similar to the query protein. The food protein sources with bioactive peptides have been screened based on the homology of protein sequences analysed using PSI BLAST Program. Bioactive peptides from albumin showing α-glucosidase and α-amylase were identified. The inhibitory activities of peptides KLPGF, EVSGL, QITKPN, AEAGVD, EAGVD, NVLQPS, LEPINF, and ANEIIF from albumin were evaluated according to the method described by Kim (Yu et al., 2012) with some modification. The peptides KLPGF and NVLQPS showed significant inhibitory effects on α-glucosidase. Peptide KLPGF, and EAGVD exhibited significant inhibitory activity against the α-amylase. Peptide KLPGF was a potential anti-diabetic inhibitor (Jakubczyk et al 2017; Wang et al., 2017).
In-silico analysis for identification of proteins with bioactive peptides
Rich source of bioactive peptides was chosen as the query sequence. A homology search was performed against the seed storage protein using PSIBLAST algorithm. The pairwise alignments of soybean glycinin with the corresponding homologues from other plant protein were stored. Using a peptide library of antioxidant and anti-hypertensive peptides, positions of these bioactive peptides were identified (Serna-Saldivar, 2015). Functional properties of protein are important in food processing and food formulation. Some of the functional properties are solubility, water and oil holding capacity, foaming capacity and stability, gelation, bulk density and viscosity. Sesame protein is used for the preparation of sweets, confectionary and bakery products (Laohakunjit et al., 2017; Nongonierma and FitzGerald, 2015).
Alpha Amylase and Alpha Glucosidase Inhibiting Peptides
During onset and development of type 2 diabetes, cellular balance of carbohydrate and lipid metabolism is affected by improper glucose metabolism (Besseiche et al., 2015). This improper regulation leads to elevated postprandial blood glucose levels. Prolonged imbalanced homeostasis for an extended time, results in hyperglycemia leading to onset of non-insulin dependent type 2 diabetes (Tang, 2015). Absorption of glucose can be delayed by reducing the rate of digestion of starch. Pancreatic alpha amylase is a key enzyme in the digestive system and catalyses the initial step in hydrolysis of starch to a mixture of smaller oligosaccharides consisting of maltose, maltotriose, and a number of α-(1-6) and α-(1-4) oligoglucans (Dhital et al., 2017; Kajaria et al., 2013). These are then acted on by α-glucosidases and further degraded to glucose which on absorption enters the blood-stream. Degradation of this dietary starch proceeds rapidly and leads to elevated PPHG (post-prandial hyperglycemia) (Chakraborty and Joy, 2017; Sacan et al., 2017). It has been shown that activity of HPA (human pancreatic α-amylase) in the small intestine correlated to an increase in post-prandial glucose levels, the control of which is therefore an important aspect in treatment of type 2 diabetes (Kajaria et al., 2013). Hence, retardation of starch digestion by inhibition of enzymes such as α-amylase plays a key role in the control of diabetes. Inhibitors of pancreatic α-amylase delay carbohydrate digestion causing a reduction in the rate of glucose absorption and lowering the post-prandial serum glucose levels (Sacan et al., 2017). An effective strategy for type 2 diabetes management is the strong inhibition of inhibition of pancreatic α-amylase (Besseiche et al, 2015; Kajaria et al., 2013). Amylase inhibitors are also known as starch blockers because they contain substances that prevent dietary starch from being absorbed by the body. Starch are complex carbohydrates that cannot be absorbed unless they are first broken down by the digestive enzyme amylase and other, secondary enzymes (Sacan et al., 2017). The presently used synthetic enzyme inhibitors cause gastrointestinal side effects such as diarrhea, flatulence, abdominal bloating etc. Therefore natural alpha amylase and glucosidase inhibitors from the dietary plants can be used as an effective therapy for treating post prandial hyperglycemia. More researchers have focused on the search for more effective inhibitors of anti-diabetic compounds from natural materials (Dhital et al., 2017), such as polysaccharides from tea leaves (Chen et al., 2014) and isoflavones from soybean (Ahmad et al., 2014; Lee et al., 2015). Egg protein hydrolysates displayed anti-diabetic activity against glucosidase activity (Lacroix and Li-Chan, 2013). Bioactive peptides from albumin showing α-glucosidase and α-amylase were identified. The inhibitory activities of peptides KLPGF, EVSGL, QITKPN, AEAGVD, EAGVD, NVLQPS, LEPINF, and ANEIIF from albumin were evaluated according to the method described by Kim (Hernández-Ledesma et al., 2004; Li-Chan, 2015) with some modification. The peptides KLPGF and NVLQPS showed significant inhibitory effects on α- glucosidase. Peptide KLPGF, and EAGVD exhibited significant inhibitory activity against the α-amylase. Peptide KLPGF was a potential anti-diabetic inhibitor (Lewis et al., 2013; Yu et al., 2012).
Antihypertensive Peptides
Hypertension is one of the major risk factors for a number of cardiovascular diseases (CVD). The risk of developing CVD is directly related to blood pressure (BP) level. It is controlled by a number of interacting biochemical pathways. Blood pressure control is partly associated with rennin angiotensin system (FitzGerald et al., 2004). Angiotensin I converting enzyme (ACE, peptidyldipeptide hydrolase) is a multifunctional ecto-enzyme that is located in different tissues and plays an important role in BP regulation (MacIsaac et al., 2014). Renin acts on angiotensinogen, inactive precursor, thus releasing decapeptide angiotensin I . ACE further removes the C-terminal dipeptide HL from angiotensin I resulting in the formation of angiotensin II, a potent vasoconstrictor (Nabi et al., 2013). ACE also removes the C-terminal dipeptide from bradykinin (potent vasodilatator) resulting in the formation of inactive peptide fragments (FitzGerald et al., 2004). ACE inhibition studies with dipeptides of varying structures, show that C-terminal tryptophan, tyrosine, phenylalanine or proline residues were most effective in enhancing substrate binding. Several ACE inhibitors contain lysine or arginine at C-terminal residue. The study has suggested that positive charge associated with side chain groups of these amino acids contribute to ACE inhibitor potency (Phelan et al., 2014; Rawendra et al., 2014). It is postulated that the mechanism of ACE inhibition involves inhibitor interaction with an anionic binding site that is distinct from the catalytic sites (Wada and Lönnerdal, 2014). This behavior is characteristic of noncompetitive inhibition. After binding to the enzyme these inhibitors deform the enzyme, and hinder binding to the substrate. This inhibition type usually occurs for enzymes containing a functional group necessary to maintain its catalytically active three-dimensional form. ACE is a zinc18 metallo enzyme (Dellafiora et al., 2015); so binding of an inhibitor to this ion could lead to a decrease in enzyme activity.
Globulin protein of sesame seeds, amandin protein of almond and cucurbitin protein of pumpkin seeds
Globulin protein was composed of acidic polypeptide (30-33 kDa) and a single basic polypeptide (20-24 kDa), isolated amandin protein was composed of acidic polypeptide (40-42 kDa) and a single basic polypeptide (15-20 kDa) and isolated cucurbitin protein was composed of acidic polypeptide (35-42 kDa) and and a basic polypeptide (20-25 kDa) (McNulty, 2006; Chhabra, 2013; Zhang et al., 2016). Amandin is an 11S globulin legumin-like protein that is hexameric in form and composed of individual polypeptides (Bojórquez-Velázquez et al., 2016). Two isoforms have been identified in almond, prunin 1 and prunin 2, which comprise native prunin (de la Cruz et al., 2017). Analyses of almond protein extract has been extensively carried out and it was found that prunin is a major component of the nut, accounting for up to 65% of the protein in the soluble extract (Zhu, 2014).
Angiotensin-converting Enzyme (ACE)
First reports of exogenous inhibitors of ACE displaying an antihypertensive effect in vivo were from snake venom (Bhat et al., 2015). These ACE inhibiting peptides had limited pharmacological application because of their lack of oral activity. Many ACE inhibiting substances have been proposed by pharmaceutical industry as drugs in treatment of hypertension such as captopril. Several side effects such as increased potassium level, reduced renal function, cough, skin, rashes, fetal abnormalities, etc., have been associated with synthetic organic drugs (Blackburn, 2017; Nawaz et al., 2017). Therefore various food proteins derived bio-functional peptides have been isolated and evaluated for their antihypertensive activity to avoid undesirable side effects of synthetic antihypertensive drugs and to avoid increasing cost of drug therapy (Fitzgerald, 2014; Manikkam et al., 2016). These peptides are mainly derived from: cheese whey, casein, zein, soyabean, fish muscle and gelatine (Puchalska et al., 2015; Sánchez and Vázquez, 2017; Shahidi and Li, 2014). The occurrence of two ACE inhibiting tetrapeptides, ALEP and VIKP in 11S globulin of amaranth seed was identified and experimentally validated by an in vitro ACE inhibition assay (Quiroga et al., 2017) (Fig. 1).
Vasodilating Effect of Nitric Oxide
Nitric oxide is made at various sites in the body and it performs important functions in many systems. Nitric oxide plays an important role in the maintenance of healthy blood pressure and, in turn, cardiovascular health (González et al., 2014). If too much nitric oxide is produced, it can dilate blood vessels dropping the blood pressure.
Mechanism of NO
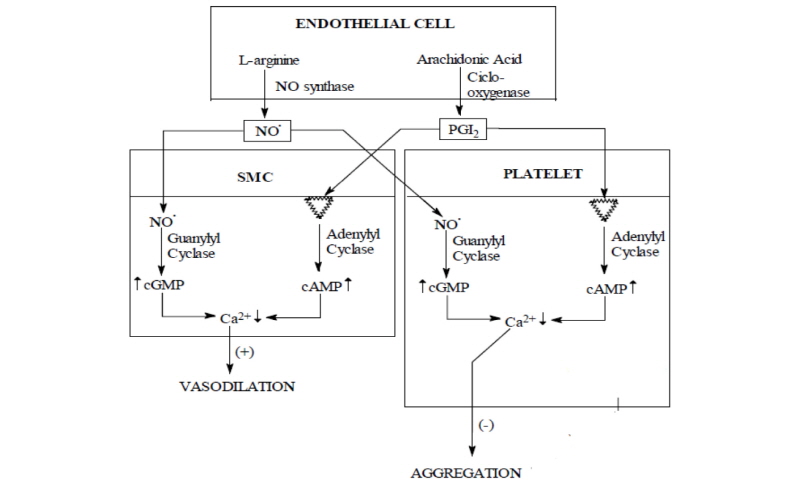
Nitric oxide is released by the endothelial cells through various stimuli, such as 5-OH-tryptamine, acetylcholine, thrombin, calcium ionophor, arachidonic acid, changes in arterial pressure, electric stimulation etc., either as NO•, or bound to a -SH group-containing carrier molecule (e.g. L-cys) that stabilizes NO release (Koch et al., 2017). Once released, NO activates the guanylate cyclase in the smooth muscle cells and platelets risings the level of intracellular messenger cGMP. This rise causes smooth muscle relaxation and platelet aggregation inhibition, presumably by a decrease in intracellular Ca2+ concentration (González et al., 2014). The endothelial vasodilator prostacyclin (PGI2) causes the same effect by the activation of adenyl-cyclase that increases the intracellular level of adenosyl monophosphate (AMP) (Fig. 2) (Luminita Jerca et al, 2002).
Spermine NONOATE and NO Donor
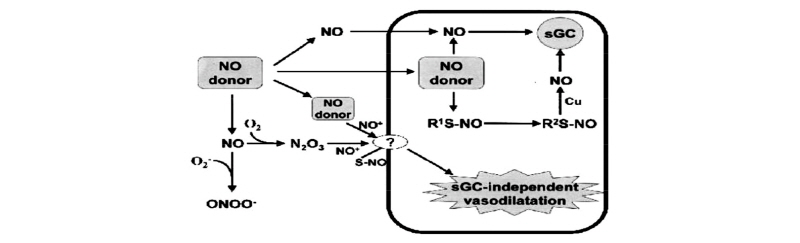
Nitric oxide (NO) is a powerful endogenous vasodilator that inhibits platelet aggregation, smooth muscle proliferation, and platelet and monocyte adhesion to the vascular wall (Koch et al., 2017). NO synthesis and/or availability is diminished in a number of cardiovascular pathologies (Quiroga et al., 2017). Delivery of exogenous NO is an attractive therapeutic option in the management of these conditions. Organic nitrates and sodium nitroprusside (SNP) are currently used for this purpose, but both have limitations. The diazeniumdiolates (NONOates) constitute a novel class of NO donor drug that decomposes spontaneously to generate up to 2 molecules of NO at physiological pH and temperature (Zhang et al., 2016). Decomposition is not catalyzed by thiols or biologic tissue, but its rate is dependent on the nucleophile adduct. (Z)- 1-{N-[3-Aminopropyl]-N-[4-(3- aminopropylammonio)butyl]-amino}-diazen-1-ium-1,2-diolate (SPER/NO) contains spermine as the nucleophile and decomposes at a relatively slow rate (Half-life = 39 minutes; 37°C; phosphate buffer, Ph 7.4) (Huang et al., 2017). SPER/NO-mediated vasodilatation and inhibition of platelet aggregation both correlate closely with the amount of NO generated. It is generally accepted that the vasodilator effects of NO are mediated via activation of smooth muscle cell soluble guanylate cyclase (sGC) to generate cyclic guanosine-3,5- monophosphate (cGMP).7 However, vasodilatation in response to high concentrations of NO is not exclusively cGMP mediated (Zhang et al., 2016). Using the selective sGC-inhibitor [1H-[1,2,4] oxadiazole [4,3- a]quinoxaline-1-one (ODQ), (Garthwaite J et al, 1995) an sGC independent component of vasodilatation has been identified in response to several NO donors (Chin et al., 2014). NONOates have also been shown to have a vasodilator effect in the presence of ODQ (Irvine et al., 2013), despite complete abolition of cGMP generation (Fig. 3) (Miller et al, 2004).
Conclusion
The current study includes the identification of bioactive peptides from sesame, almond and pumpkin seed protein. The 11S globulin, amandin and cucurbitin protein are the major seed storage proteins, dissociated into acidic and basic polypeptides. This indicated the presence of disulphide bond between the acidic and basic polypeptide. Homology searches revealed the presence of antihypertensive and alpha amylase inhibiting peptides in 11S globulin, amandin and cucurbitin proteins. 11S globulin protein of sesame seed was confirmed by MALDI-TOF. Alpha amylase inhibiting activities of proteins were evaluated and it was found that the cucurbitin protein has higher inhibiting activity.
Future Perspectives
Research targeted towards fabrication, characterization, and efficacy in testing towards antihypertensive peptides continues to evolve promptly. Recent efforts include the practice of in-silico methods to perform virtual enzyme hydrolysis of proteins for which the released peptide sequences can be synthesized and tested for activity. If in vitro activity (usually against renin and ACE) is confirmed, actual enzyme hydrolysis can be performed to obtain peptides that can then undergo in vitro and in vivo testing. Hence in silico approach could be act as less time-consuming and inexpensive approach whereby various enzymes are thrown at proteins followed by testing of each hydrolysate to identify active products. However, availability of the primary structure of several more proteins is required to expand the in silico method beyond the currently limited structurally characterized food proteins. In addition the structure-and functional properties of antihypertensive peptides developed using improved enzyme digestion tools that will release targeted peptides and some untargeted peptides. Finally, more human intermediation trials required for to claim of antihypertensive activity on peptide-formulated food products or nutraceuticals.
