

서 론
재료 및 방법
실험재료
세포 종류 및 시약
SH-SY5Y 세포주 배양
세포 생존율 측정
Western blot을 통한 단백질 발현 측정
통계분석
연구결과 및 고찰
Kaempferol, quercetin 및 그 배당체의 H2O2에 대한 세포생존율 개선 효과
Kaempferol, quercetin 및 그 배당체의 세포사멸 관련 단백질 발현 조절 효과
결 론
서 론
전 세계적으로 인구의 고령화에 따라 치매와 같은 신경퇴행성 질환의 발병률이 높은 실정이며, 뇌에서 산화적 스트레스(oxidative stress)는 신경퇴행성 질환의 대표적인 원인으로 알려져 있다(Albers and Beal, 2000). Reactive oxygen species(ROS)는 superoxide radical, hydroxyl radical, hydrogen peroxide(H2O2) 등의 다양한 종류를 포함하며, 체내에서 이들의 과다 축적 시 산화적 손상을 일으켜 세포막 손상, 단백질 산화, DNA 변성 등을 초래한다(Apel and Hirt, 2004; Valko et al., 2007). 특히, 뇌에서 산화적 스트레스가 유발되었을 때, 염증반응, 시냅스 손상, mitochondria dysfunction 등을 일으켜 세포사멸에 관여하는 여러 인자 발현을 변화시킨다(Coimbra-Costa et al., 2017). 산화적 자극을 받은 신경세포의 mitochondria는 cytochrome C를 방출하여 caspase-9, caspase-3, poly(ADP-ribose) polymerase 단백질의 cleavage를 유도시켜 세포사멸을 유도한다(Coimbra-Costa et al., 2017; Eldadah and Faden, 2000). 또한, 산화적 손상은 세포사멸을 억제하는 B-cell lymphoma 2(Bcl-2) 단백질 발현을 감소시키고, 세포사멸을 유도하는 Bcl-2-associated X protein(Bax) 단백질의 발현을 증가시켜 신경세포 사멸에 관여하는 것으로 알려져 있다(Cai and Shen, 2018). 따라서 세포사멸 기전 조절을 통해 신경세포의 산화적 손상에 대한 보호 효과를 나타낼 수 있는 다양한 천연식물 및 생리활성 성분들의 개발에 관한 연구가 활발히 이루어지고 있다(Zhao, 2009).
최근 많은 연구들은 산화적 스트레스가 신경퇴행성 질환의 주요 원인이라고 보고하였으며, 이에 따라 천연 항산화제를 신경퇴행성 질환의 치료에 이용하고자 하는 연구가 활발히 이루어지고 있다(Albers and Beal, 2000; Zhao, 2009). 특히, 신경퇴행성 질환 예방 및 치료를 위한 소재 개발을 위해, H2O2 등의 ROS의 처리로 산화적 스트레스가 유도된 신경세포 모델이 널리 사용되고 있다(Zheng et al., 2019). H2O2는 산화적 스트레스를 유도하는 대표적인 ROS의 일종으로, 뇌에서 반응성이 높은 hydroxyl radical을 생성하여 신경세포 손상을 일으킨다(Rhee, 1999). 뿐만 아니라, H2O2는 신경퇴행성질환의 뇌에서 염증반응, 세포사멸을 유도하는 것으로 보고되었다(Li et al., 2003).
Flavonoid는 노란색 또는 담황색을 나타내는 polyphenol 화합물의 일종으로, 과일 및 엽채류 등의 식물계에 풍부하게 분포되어 있으며, 대부분 당과 결합된 배당체(glycoside)의 형태로 존재한다(Miean and Mohamed, 2001). Flavonoid는 산화적 스트레스를 유발하는 free radical, ROS 등을 효과적으로 억제하여 항산화 활성이 우수한 생리활성물질로 알려져 있다(Zhao, 2009). Kaempferol과 quercetin은 식물계에 풍부하게 존재하는 flavonoid의 일종으로써, 이들의 생리활성에 관한 연구가 활발히 진행되고 있다(Miean and Mohamed, 2001). 특히 신경퇴행성 질환의 일종인 알츠하이머성 치매 환자의 증상 완화 및 개선 효능이 보고된 은행나무(Ginkgo biloba), 차나무(Camellia sinensis), 감초(Glycyrrhiza uralensis) 등 식물체에 kaempferol 및 quercetin이 풍부하게 함유되어있어, 이들 flavonoid가 뇌 기능 개선에 중요한 역할을 할 것으로 보고되었다(Alexander et al., 2023). 이외에도, kaempferol 및 quercetin은 free radical 생성 억제, ROS 소거능, 지질과산화 억제, 항산화 효소 활성 증가 등의 항산화 작용으로 인해 항암, 항당뇨, 지질대사 개선, 산화적 손상으로 인한 신장, 간 손상 개선 효과 등 다양한 질병에 대한 효능이 보고되었다(Du et al., 2018; Hou et al., 2004; Kamada et al., 2005; Roslan et al., 2017). 사전 연구를 통해 머위(Petasites japonicus) 추출 및 분획물 중 ethyl acetate(EtOAc) 분획물이 다른 추출 및 분획물에 비해 우수한 항산화 활성과 신경교세포 보호 효과를 나타냄을 확인하였으며, EtOAc 분획물에는 kaempferol과 quercetin에 glucoside가 결합된 형태의 배당체가 주요 활성물질임을 확인하였다(Lee et al., 2015; Wang et al., 2014). 자연적으로 flavonoid는 식물체 내에 당과 결합된 형태의 배당체 형태로 주로 존재하는 것으로 알려져 있다(Xiao, 2017). Kaempferol과 quercetin은 다양한 자극에 의한 뇌 손상에 대해 항산화, 항염증 작용 등을 통한 신경보호 효과 등이 보고되었으나(Wang and Liu, 2018; Yang et al., 2014), kaempferol과 quercetin 및 그 배당체의 H2O2로 유도된 산화적 스트레스에 대한 신경세포 보호 효과 비교에 관한 연구는 부족한 실정이다.
본 연구에서는 사람의 신경모세포종인 SH-SY5Y 신경세포를 이용하여 flavonoid의 일종인 kaempferol, quercetin과 그 배당체인 kaempferol-3-O-glucoside와 quercetin-3-β-D-glucoside의 H2O2 유도 산화적 스트레스에 대한 신경세포 보호 효과를 살펴보고자 하였다. 또한, 신경세포 보호 작용 기전 확인을 위해, 세포사멸에 관여하는 단백질 발현 측정을 통해 flavonoid의 신경세포 보호 메커니즘을 규명하고자 하였다.
재료 및 방법
실험재료
본 실험에 사용한 kaempferol, quercetin, quercetin-3-β-D-glucoside는 Sigma(Sigma, St. Louis, MO, USA)사에서, kaempferol-3-O-glucoside는 Extrasynthese(Genay, France)사에서 구입하였으며 각 구조는 Fig. 1에 제시하였다.

세포 종류 및 시약
실험에 사용한 SH-SY5Y 신경세포는 한국세포주은행(Seoul, Korea)에서 분양받아 실험에 사용하였다. 세포 배양을 위한 Dulbecco’s modified eagle medium(DMEM), fetal bovine serum(FBS), penicillin-streptomycin, trypsin-EDTA 시약은 Welgene(Gyeongsan, Korea)에서 구입하여 사용하였다. 세포의 산화적 스트레스 유도를 위한 H2O2는 Junsei(Tokyo, Japan)사 제품을 사용하였다.
SH-SY5Y 세포주 배양
SH-SY5Y 세포는 10% FBS, 100 units/mL의 penicillin-streptomycin을 함유한 DMEM을 이용하여 37°C, 5% CO2 incubator에서 배양하였다. 2-3일에 한 번씩 배지를 교환하면서 세포를 배양하였으며, 세포 증식이 최대에 도달하였을 때 0.05% trypsin과 0.02% EDTA 혼합액을 이용하여 계대배양 하면서 실험에 사용하였다.
세포 생존율 측정
세포 생존율은 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide(MTT) assay를 이용하여 측정하였다(Mosmann, 1983). 세포 증식이 80-90%에 도달하였을 때, 세포를 5 × 104 cells/mL로 plate에 seeding하여 세포를 부착시킨 후, 각 시료를 0.5, 0.25, 1 µM의 농도별로 처리하여 37°C, 5% CO2 incubator에서 4시간 배양시켰다. 그 뒤, 세포의 산화적 손상 유도를 위해 300 µM H2O2를 처리하여 24시간 배양시켰다. 세포 생존율 측정을 위해 5 mg/mL MTT solution(Bio Basic, Toronto, Canada)을 처리하여 4시간 배양한 뒤, 환원된 formazan 결정을 dimethyl sulfoxide(Bio pure, Ontario, Canada)에 용해시켜 540 nm에서 흡광도를 측정하였다. 세포 생존율은 normal군의 생존율에 대비하여 %로 나타내었다.
Western blot을 통한 단백질 발현 측정
세포를 1 × 106 cells/mL로 seeding하여 37°C, 5% CO2 incubator에서 24시간 배양시킨 뒤, 각 시료를 1 µM의 농도로 처리하여 4시간 재배양시켰다. 그 뒤, 세포 손상을 유도하기 위해 H2O2를 300 µM의 농도로 처리하여 24시간 배양시켰다. 배양한 세포는 protease inhibitor cocktail이 포함된 RIPA buffer(Elpis Biotechnology, Daejeon, Korea)에 의해 용해시킨 뒤, Bio-Rad protein assay kit(Bio-Rad, Hercules, USA)를 이용하여 단백질을 정량하였다. 각 군별 동량의 단백질 시료 15 µg을 10-13%의 sodium dodecyl sulphate-polyacrylamide gel(SDS-PAGE)에서 전기영동한 뒤, polyvinylidene fluoride membrane(Millipore, Billerica, MA, USA)에 transfer하였다. 그 뒤, blocking을 위해 membrane을 5% skim milk를 이용하여 실온에서 1시간 배양시켰다. Blocking한 membrane은 1차 항체인 caspase-3(#9662, Cell Signaling Technology, Beverly, MA, USA), Bax(#sc-493, Santa Cruz, MA, USA), Bcl-2(#sc-492, Santa Cruz, MA, USA) 단백질과 4°C에서 overnight 반응시킨 뒤, 2차 항체(Cell Signaling Technology, Beverly, MA, USA)와 실온에서 1시간 반응시켰다. 단백질 발현은 enhanced chemiluminescence solution(Bio-Rad, Hercules, USA)을 가하여 발광시켜, chemiluminescence image system(Davinch-ChemiTM, Davinch-K Co, Seoul, Korea)을 이용하여 확인하였다.
통계분석
각 실험 결과는 평균 ± 표준편차로 나타내었고, 통계분석은 Statistical Package for the Social Science(SPSS) software를 이용하여 수행하였다. 각 그룹간의 결과를 비교하기 위해서 one-way analysis of variance(ANOVA)를 구한 후, Duncan’s multiple range test(P < 0.05)를 실시하여 유의성을 검증하였다.
연구결과 및 고찰
Kaempferol, quercetin 및 그 배당체의 H2O2에 대한 세포생존율 개선 효과
Kaempferol, quercetin과 그 배당체인 kaempferol-3-O-glucoside, quercetin-3-β-D-glucoside의 H2O2로 산화적 손상이 유발된 SH-SY5Y 신경세포에서 보호 효과를 세포 생존율 측정을 통해 살펴보았다. 먼저 시료가 SH-SY5Y 신경세포에 미치는 독성을 확인하기 위해 4가지 flavonoids를 농도별로 처리하여 세포생존율을 측정한 결과, 최대 1 µM의 농도에서 세포 생존율에 영향을 미치지 않음을 확인하여(data not shown), 0.25, 0.5, 1 µM의 농도에서 H2O2에 대한 신경세포 보호 효과를 확인하였다. 아무것도 처리하지 않은 normal군의 세포 생존율을 100% 라고 했을 때, H2O2만을 처리한 control군의 경우 65.32%의 낮은 세포 생존율 나타내어 H2O2 처리로 인한 신경세포의 산화적 손상을 확인하였다. 반면, kaempferol 및 kaempferol-3-O-glucoside를 처리하였을 경우(Table 1), control군에 비해 유의적인 세포 생존율의 증가를 확인하였다. 특히 kaempferol을 처리한 군은 모든 농도에서, kaempferol-3-O-glucoside는 0.5, 1 µM의 농도로 처리한 군에서 70% 이상의 세포 생존율 수치를 나타내어, kaempferol과 kaempferol-3-O-glucoside의 산화적 손상에 대한 신경세포 보호 효과를 확인하였다. 또한, quercetin과 quercetin-3-β-D-glucoside를 처리하였을 때(Table 2), 모든 농도에서 70% 이상의 생존율을 나타내었으며, 특히 quercetin을 처리하였을 경우 농도 의존적으로 세포 생존율이 증가하였으며, 1 µM의 농도에서 94.37%의 우수한 생존율을 나타내었다. 따라서 4가지 flavonoid는 H2O2로 처리로 인해 산화적 스트레스가 유도된 신경세포에서의 세포 생존율 개선을 통해 신경세포 보호 효과를 확인할 수 있었다.
Table 1.
Effect of kaempferol and kaempferol-3-O-glucoside on cell viability of SH-SY5Y cells treated with H2O2
| Treatment (µM) | Cell viability (%) | |
| Normal | 100.00 ± 1.38a | |
| H2O2-treated control | 65.32 ± 2.80d | |
| H2O2 + Kaempferol | 0.25 | 73.79 ± 0.38b |
| 0.5 | 74.27 ± 2.03b | |
| 1.0 | 71.56 ± 1.67b | |
| H2O2 + Kaempferol-3-O -glucoside | 0.25 | 68.41 ± 1.79c |
| 0.5 | 74.44 ± 1.87b | |
| 1.0 | 71.40 ± 2.88b | |
Table 2.
Effect of quercetin and quercetin-3-β-D-glucoside on cell viability of SH-SY5Y cells treated with H2O2
| Treatment (µM) | Cell viability (%) | |
| Normal | 100.00 ± 1.38a | |
| H2O2-treated control | 65.32 ± 2.80f | |
| H2O2 + Quercetin | 0.25 | 73.27 ± 0.58de |
| 0.5 | 76.59 ± 2.07cd | |
| 1.0 | 94.37 ± 2.56b | |
|
H2O2 + Quercetin-3-β-D -glucoside | 0.25 | 77.04 ± 1.69c |
| 0.5 | 72.37 ± 3.59e | |
| 1.0 | 75.70 ± 1.69cde | |
이전 연구에 의하면 SH-SY5Y 신경세포에 H2O2의 처리 시, 세포의 산화적 손상으로 인해 세포 생존율이 감소하는 것으로 보고되었다(Suthprasertporn et al., 2019). 본 연구에서도 H2O2만을 처리한 control군에서 세포 생존율이 감소하여 산화적 스트레스로 인한 SH-SY5Y 신경세포의 손상을 확인하였다. 또한, 본 연구에서 4가지 flavonoid를 각각 0.25, 0.5, 1 µM의 농도로 처리 했을 때 모든 농도에서 유의적으로 세포생존율 수치가 증가하여 신경세포 보호 효능을 나타내었으며, 특히 quercetin 1 µM의 처리 시 94.37%의 높은 세포 생존율 수치를 확인하였다. Kaempferol과 quercetin은 항산화 활성이 우수한 것으로 알려져, 이들의 신경퇴행성 질환 소재로써의 가능성에 관한 연구도 활발히 이루어지고 있다(Dai et al., 2018; Hussein et al., 2018; Kouhestani et al., 2018; Wang and Liu, 2018). Kaempferol은 산화적 손상이 유도된 동물의 뇌 조직에서 GSK3β, Nrf2 pathway 조절과 항산화 효소 활성을 증가시켜 산화적 손상을 개선 시켰을 뿐만 아니라, 염증성 cytokine 억제를 통한 항염증 효능을 나타내어 손상된 뇌에서의 보호 효과가 보고되었다(Hussein et al., 2018; Kouhestani et al., 2018). Quercetin과 그 배당체인 quercetin-3-β-D-glucoside는 산화적 손상이 유도된 뇌 조직에서 ROS 소거능, malondialdehyde와 같은 산화적 손상과 관련된 factor를 억제하였으며, superoxide dismutase, catalase와 같은 항산화 효소 활성을 증가시켜 뇌에서 항산화 활성이 우수한 것으로 확인되었다(Dai et al., 2018; Wang and Liu, 2018). 그러나 kaempferol, kaempferol-3-O-glucoside, quercetin, quercetin-3-β-D-glucoside와 같은 4가지 flavonoid의 산화적 손상에 대한 신경세포 보호 효과 비교에 관한 연구는 부족한 실정이며, 따라서 본 연구에서 4가지 flavonoid의 H2O2 유도 산화적 스트레스에 대한 신경세포 보호 효과와 세포 사멸 관련 기전 확인을 통해 신경세포 보호 메커니즘을 규명할 수 있었다. Flavonoid 기본 구조에 당이 붙지 않은 형태인 kaempferol 및 quercetin 등 aglycone 형태는 지용성의 특징을 갖는 반면 식물체에서는 주로 당과 결합된 배당체 형태로 존재하며 이는 친수성의 특징을 갖는다(Jiang et al., 2015). 이전 연구에 의하면, quercetin은 quercetin 배당체에 비해 뇌의 blood-brain barrier(BBB) 투과성이 증가되어 신경 보호 효과도 높은 것으로 보고되었다(Ossola et al., 2009). 또한 quercetin은 quercetin 배당체 형태에 비해 지질과산화 억제 효과가 높아 항산화 활성이 우수한 것으로 보고되었다(Ratty and Das, 1988). 본 연구에서도 quercetin이 그 배당체에 비해 세포생존율 개선 효과가 우수한 것으로 보고되었는데, 사전 보고된 연구와 유사한 경향을 나타냄을 알 수 있었다. 이전 연구에서 25 µM의 kaempferol을 glutamate로 산화적 손상이 유도된 hippocampal neuronal cells에 처리 시, calcium influx 조절, ROS 소거능, lactate dehydrogenase 활성 조절 등에 의해 세포 생존율의 증가를 확인하였다(Yang et al., 2014). 또한, kaempferol 3-O-beta-rutinoside, 6-hydroxykaempferol 3,6-di-O-beta-D-glucoside와 같은 kaempferol 유도체는 산화적 손상이 유도된 신경세포에서 세포 생존율 개선을 나타내었다(Qu et al., 2009). 산화적 스트레스가 유도된 신경세포에서 quercetin 및 그 배당체인 quercetin-3-β-D-glucoside을 처리했을 때, 세포 생존율 증가를 통해 신경보호 효과가 보고되었고, 이는 ROS와 지질과산화 생성의 감소 및 항산화 효소 활성 증가로 인한 것임을 확인하였다(Bao et al., 2017; Dai et al., 2018; Jembrek et al., 2012). 따라서 이전 연구 논문들과 본 연구결과를 종합해 볼 때, 4가지 flavonoid는 산화적 손상이 유도된 신경세포에서 세포 생존율 개선을 통해 신경세포 보호 효능이 있는 것으로 생각된다.
Kaempferol, quercetin 및 그 배당체의 세포사멸 관련 단백질 발현 조절 효과
Caspase protease는 세포가 정상적인 상태에서 핵과 mitochondria의 외막에 불활성 형태로 존재하지만, 산화적 손상 시 활성화되어 DNA cleavage 등에 인해 많은 기질 단백질을 분해하여 세포사멸를 유도한다(Singhal et al., 2005). 이 중 caspase-3는 세포사멸에 관여하는 많은 단백질의 분절을 촉진하는 사멸 단백분해효소로써, DNA repair, DNA stability, 전사 조절에 관여하는 PARP 단백질을 분해시키는 것으로 알려져 있다(Muller et al., 1994). 또한, 세포사멸에 관여하는 인자인 Bcl-2 family는 mitochondria 외막에 존재하여 mitocondria의 보존과 세포사멸을 조절하는 단백질로, 세포사멸를 억제하는 Bcl-2, Bcl-xl, Bcl-b 등과 세포사멸를 유발하는데 관여하는 Bax, Bak, Bid 등으로 구성되어있다(Nikiforos and Kontogeorgos, 2000). 이들의 균형이 깨어질 시 mitochondria 내부에서 cytosol로 cytochrome C가 방출되어 caspase, p53 등의 활성으로 apoptosis가 유발되는 것으로 알려져 있다(Chang et al., 2000). 이전 연구에서 SH-SY5Y 신경세포에 H2O2의 처리로 산화적 손상을 유도하였을 때, 세포사멸 유발인자인 caspase-3가 활성화되어, cleavage 형태인 cleaved caspase-3의 발현이 증가하여 세포사멸이 유도되었음을 확인하였다(Lee et al., 2018; Park et al., 2015). 뿐만 아니라, SH-SY5Y 신경세포에서의 산화적 손상은 세포사멸에 관여하는 Bcl-2 family 조절을 통해 세포사멸을 유도하는 것으로 보고되었다(Lee et al., 2018).
본 연구에서 4가지 flavonoid의 산화적 손상에 대한 신경세포 보호 작용 기전을 확인하기 위해, 세포사멸에 관여하는 것으로 알려진 caspase-3, Bax, Bcl-2 단백질 발현을 확인하였다. 본 연구에서 4가지 flavonoid를 각각 1 µM의 농도로 처리한 뒤, H2O2로 산화적 손상을 유도한 SH-SY5Y 신경세포의 세포사멸 관련 단백질 발현을 측정하였다. Normal군에 비해 H2O2로 산화적 손상을 유도한 control군에서 cleaved caspase-3, Bax와 같은 세포사멸을 유도하는 단백질 발현의 증가를 확인하였으며, 세포사멸을 보호하는 Bcl-2 단백질 발현의 감소를 확인하여 세포사멸로 인한 SH-SY5Y 신경세포 손상을 확인하였다. 반면, kaempferol을 처리한 군에서는 세포사멸 관련 단백질 발현의 유의한 차이를 확인하지 못했으나, 그 배당체인 kaempferol-3-O-glucoside를 처리하였을 경우 Bax 단백질 발현의 감소를 통해 세포사멸에 대한 보호 효과를 확인하였다(Fig. 2). 또한 quercetin과 그 배당체인 quercetin-3-β-D-glucoside의 세포사멸 조절 기전을 확인한 결과(Fig. 3), quercetin과 quercetin-3-β-D-glucoside는 control군에 비해 세포사멸 유발인자인 cleaved caspase-3, Bax 단백질을 유의적으로 감소시켜 항세포사멸 효과가 있는 것으로 사료된다. 따라서 kaempferol-3-O-glucoside, quercetin, quercetin-3-β-D-glucoside을 처리한 군에서 caspase 3 및 Bax 단백질 발현 억제를 통해 세포사멸 기전을 조절함으로써 신경세포의 보호 효과를 나타내는 것을 확인할 수 있었다.
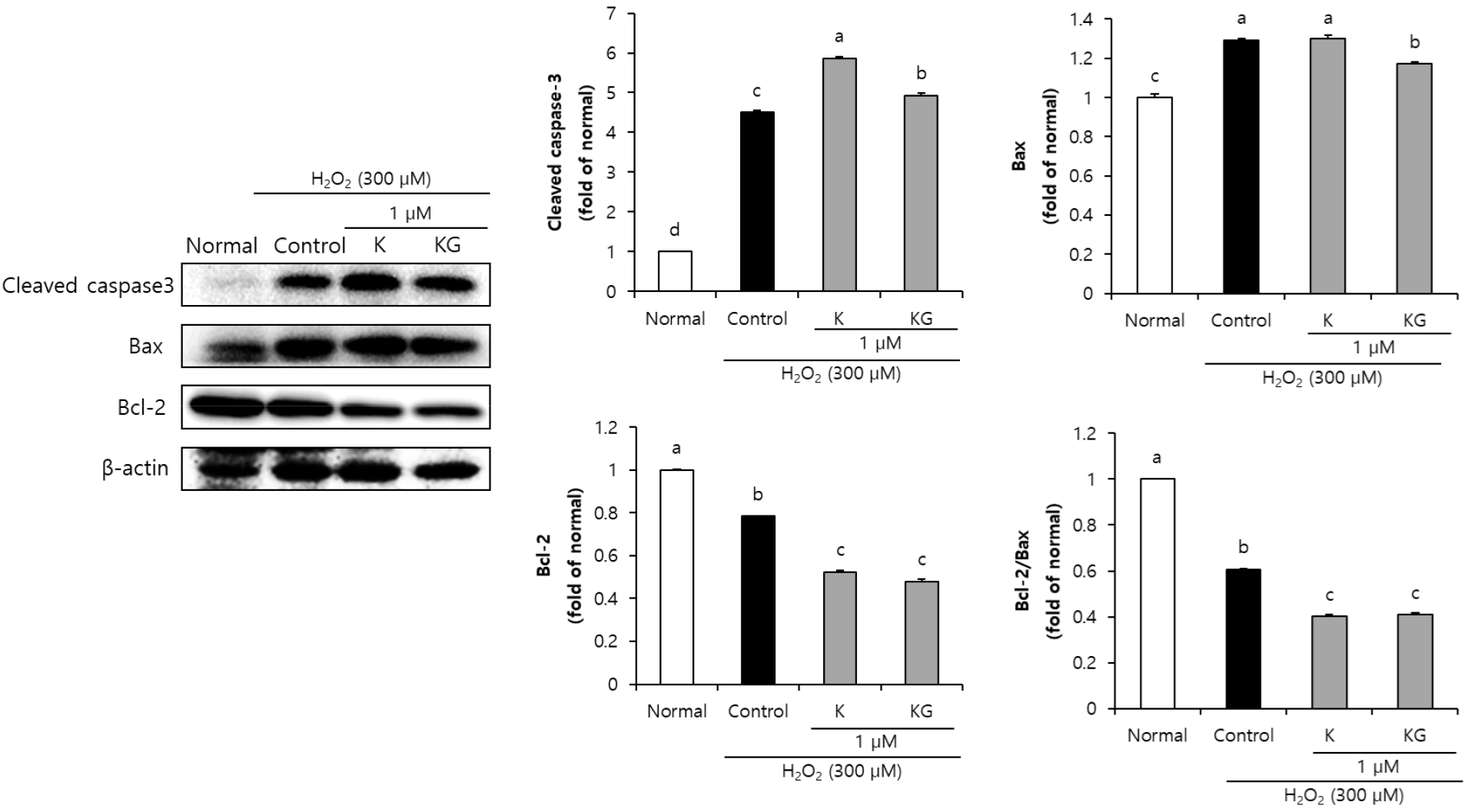
Fig. 2.
Effect of kaempferol and kaempferol-3-O-glucoside on apoptosis-related protein expression levels of SH-SY5Y cells treated with H2O2.
Values are means ± SD. a-dMeans with different letters are significantly different (P < 0.05) according to Duncan’s multiple range test. β-Actin was used as a loading control. K, kaempferol; KG, kaempferol-3-O-glucoside; Q, quercetin; QG, quercetin-3-β-D-glucoside.
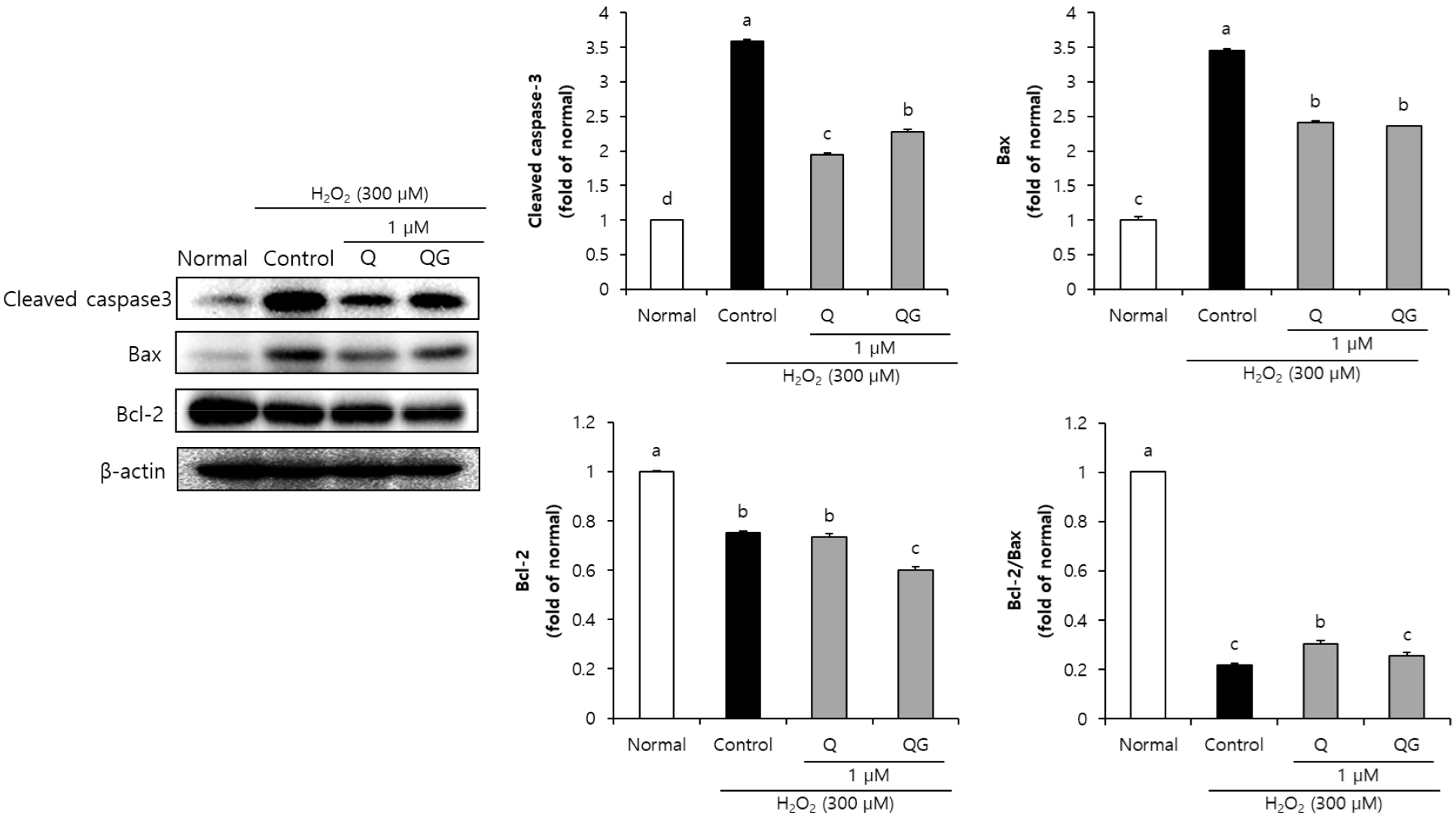
Fig. 3.
Effect of quercetin and quercetin-3-β-D-glucoside on apoptosis-related protein expression levels of SH-SY5Y cells treated with H2O2.
Values are means ± SD. a-dMeans with different letters are significantly different (P < 0.05) according to Duncan’s multiple range test. β-Actin was used as a loading control. K, kaempferol; KG, kaempferol-3-O-glucoside; Q, quercetin; QG, quercetin-3-β-D-glucoside.
Flavonoid는 하나의 heterocyclic ring에 결합된 2개의 phenyl ring의 기본구조(C6-C3-C6)를 갖는다(Shen et al., 2022). Kaempferol과 quercetin은 동일한 3-hydroxyl flavone 기본구조로 구성되어 있으나, quercetin은 kaempferol에 비해 phenyl ring에 hydroxyl 작용기가 하나 더 결합된 구조이다(Dabeek and Marra, 2019). 이전 연구에 의하면, flavonoid 화합물의 hydroxyl 작용기 개수는 해당 화합물의 생리활성에 밀접한 영향을 미치는 것으로 보고되어있다(Kumar and Pandey, 2013). 본 연구에서 quercetin이 kaempferol에 비해 우수한 세포사멸 관련 인자 조절 효과를 나타낸 것은 이러한 구조적 차이 때문일 것으로 생각된다. Kaempferol 처리 시, 본 연구에서의 cleaved caspase-3, Bax 및 Bcl-2 세포사멸 조절 기전에 control군에 비해 유의한 효과를 나타내지 못했지만 산화적 손상을 유도한 신경세포에서 Bid, MAPK와 같은 인자 발현 조절을 통해 세포사멸 보호 효과를 나타냄이 보고된 바 있다(Yang et al., 2014). 본 연구에서 quercetin과 quercetin 배당체를 처리한 결과에서는 aglycone과 배당체 두 화합물 모두에서 세포사멸 유도 인자인 cleaved caspase-3 및 Bax 발현 감소를 나타내었다. Bao et al.(2017)의 연구에 의하면, 산화적 손상이 유도된 PC-12 신경세포에서 quercetin은 세포사멸 유도인자인 Bax, cleaved caspase-3 단백질 발현 감소와 항세포사멸 인자인 Bcl-2 단백질 증가를 확인하여 본 연구와 유사한 결과 나타냄을 알 수 있었다. 또한, quercetin-3-β-D-glucoside는 산화적 손상이 유도된 뇌에서 cleaved caspase-3 단백질 발현 감소와 Bcl-2 단백질 발현 증가를 확인하여 세포사멸 조절을 통한 신경보호 효과가 보고된 바 있다(Dai et al., 2018). 따라서, 4가지 flavonoid는 세포사멸 관련 인자 조절을 통해 신경세포 보호 효과를 나타내는 것으로 사료된다.
본 연구에서는 kaempferol, quercetin과 그 배당체인 kaempferol-3-O-glucoside와 quercetin-3-β-D-glucoside의 H2O2의 처리로 산화적 손상이 유도된 SH-SY5Y 신경세포에서 세포사멸에 관여하는 단백질 발현 조절을 통해 신경세포 보호 효능을 확인하였다. 이들 flavonoid 중에서 quercetin의 처리 시 다른 flavonoid에 비해 높은 세포생존율을 나타내어 우수한 신경세포 보호 효과를 나타내었다. 또한 kaempferol을 제외한 나머지 flavonoid는 cleaved caspase-3 및 Bax 단백질 발현 억제를 통해 신경세포 보호 효과를 나타내는 것을 알 수 있었다. 본 연구는 kaempferol, quercetin 및 그 배당체의 cellular system에서 신경세포 보호 효과를 비교한 연구로서, 배당체 및 비배당체의 소화, 흡수 및 대사과정에서 생체이용률을 고려한 신경 보호 효능에 대한 한계가 있다. 따라서 추후 in vivo에서의 이들 flavonoid의 신경 보호 관련 연구가 필요하며, 세포 내 ROS 생성 억제능, 항산화 효소 활성, 유세포 분석 등의 추가 실험을 통해 보다 구체적인 신경세포 보호 메커니즘을 규명함으로써, 신경퇴행성 질환의 예방 및 치료 소재로서의 가능성을 더욱 명확히 제시할 수 있을 것으로 생각된다.
결 론
뇌에서 산화적 스트레스는 알츠하이머 질환과 같은 신경퇴행성 질환의 원인으로 알려져 있다. 본 연구에서는 flavonoid의 일종인 kaempferol, quercetin, kaempferol-3-O-glucoside, quercetin-3-β-D-glucoside의 H2O2 유도 산화적 손상에 대한 SH-SY5Y 신경세포 보호 효과에 대해 확인해보았다. H2O2의 처리로 산화적 손상을 유도한 신경세포에서 4가지 flavonoid를 각각 처리 시, control군에 비해 세포생존율 증가를 확인하여 flavonoid의 신경세포 보호 효과를 알 수 있었다. 특히, 1 µM quercetin의 처리 시 94.37%의 높은 세포 생존율 수치를 나타내어, 다른 flavonoid에 비해 우수한 신경세포 보호 효과를 알 수 있었다. 신경세포 보호 작용 메커니즘을 확인하기 위해 세포사멸 관련 인자인 cleaved caspase-3, B-cell lymphoma 2(Bcl-2), BCl-2-associated X protein(Bax) 등의 단백질 발현을 측정하였다. kaempferol-3-O-glucoside를 처리한 경우 Bax 단백질 발현 감소를 통해 세포사멸 조절 효과를 확인하였다. 또한 quercetin과 quercetin-3-β-D-glucoside를 처리한 군의 경우 세포사멸 유도 인자인 cleaved caspase-3, Bax 단백질 발현 감소를 통해 세포사멸 조절 기전을 확인하였다. 본 연구에서는 4가지 flavonoid의 세포사멸 인자 조절을 통한 신경세포에서 산화적 스트레스 개선 효과를 확인하였으며, 본 연구를 통해 이들 flavonoid의 신경퇴행성 질환 치료 소재로써의 가능성을 제시하고 한다.
