

Introduction
Materials and Methods
Plant materials and growing conditions
Treatment
Analysis of growth
High-Performance Liquid Chromatography (HPLC) for centellosides analysis
Statistical analysis
Results and Discussions
Plant growth parameters
Centelloside content
Correlation between growth parameters and triterpenoids
Conclusion
Introduction
Centella asiatica is a perennial medicinal herb that possesses large quantities of pentacyclic triterpenoid saponins and has been used in traditional medicine (James and Dubery, 2009). Centelloside, such as asiaticoside, madecassoside, asiatic acid, and madecassic acid, are major bioactive triterpenoids of C. asiatica (Müller et al., 2013). They exhibit numerous therapeutic properties, beneficial for treating diseases associated with the blood, skin, and nervous system (Bonfill et al., 2011).
The effectiveness of bioactive compounds hinges on the concentration of bioactive molecules in medicinal plants. Furthermore, controlled abiotic stresses can be utilized to enhance the nutraceutical value. It is essential to comprehend how the cultivation process and stress conditions influence the biosynthesis of these molecules. However, the large-scale cultivation of C. asiatica remains significantly underexplored. As a result, a large portion of industrial demand is satisfied by noncommercial farmers and wild-grown centella, leading to variations in the compositions and concentrations of centelloside due to differing growth conditions (Thomas et al., 2010).
Light is a critical abiotic factor for plants, and its intensity, duration, and wavelength significantly impact the photosynthesis process, metabolism, and morphological development of plants (Choi et al., 2022). In indoor farming, crop production solely relies on artificial lighting, offering the advantage of regulating light units for enhanced yield. On the other hand, indoor farming commonly practiced by hydroponic system. This system is preferable due to the ability of controlling the root zone environment of plants. As well, the growth and development of roots depend on adequate carbohydrate supply from photosynthesis byproducts or stored reserves. Therefore, to ensure root growth of plant, proper management of aboveground environmental factors including light intensity is required. Thus, efficient light management is essential for profitable returns. Currently, light-emitting diodes (LEDs) represent a promising technology with significant advantages, including high photoelectric conversion efficiency (Bian et al., 2017). This study investigated the morphological growth of C asiatica and we want to evaluate the impact of light on its centelloside content.
Mass cultivation of Centella asiatica is commercially inadequate, and existing supply relies on low-quality from the wild. However, clean cultivation of C. asiatica with good quality is desired by the pharmaceutical industries. Considering the above reasons, indoor farming using the hydroponic system is an opportunity to cleanly cultivate C. asiatica and scope to meet the present medicinal demand. Currently, some research has been conducted on light intensity and accumulation of bioactive substances in Centella asiatica, but research on metabolite changes in hydroponic cultivation systems using LED lighting is lacking (Nawae et al., 2021; Song et al. 2022). Therefore, the purpose of this study is to fill this gap and improve the possibility of industrial use. This study is expected to contribute to increasing the indoor cultivation and productivity of high-quality Centella asiatica by systematically analyzing changes in the growth and their centelloside accumulation according to light intensity.
Materials and Methods
Plant materials and growing conditions
One-month-old seedlings of Centella asiatica (L.) were collected from Hapcheon, Gyeongnam, Korea. Multiplication of seedlings was carried out through vegetative propagation via stolon in a seedling tray (external dimensions: width 540 mm × length 280 mm; 50 holes) using commercial growing media from Seoul Bio Co., Ltd., Eumseong-gun, Korea. The study was conducted in an environmentally controlled room (temperature 25°C, humidity 65%). The nutrient film technique (NFT) was used in a hydroponic system. For acclimatization, propagated plantlets were transplanted into a small growing tank filled with a hydroponic solution (formula provided by the University of Seoul, Shawon et al., 2023) for 7 days. The electrical conductivity (EC) of the solution was 0.2 dSm-1 for the first three days, increasing to 0.6 thereafter. Uniformly sized plantlets (height 5 cm, with 4 to 5 leaves each) were transferred to a tank (46 cm × 32 cm × 24 cm; KSP-1000, Gahwa Tech, Korea) equipped with NFT for the experiment. A schematic diagram of this NFT system was shown in our previous study (Shawon et al., 2023). The upper section of the tank supports the plants and ensures nutrient distribution to roots, while the lower section stores additional solution. A total of 15L of nutrient solution was circulated from the lower to the upper section of the tank by an underwater motor, and then back to the lower section through a specific hole.
The room temperature and relative humidity were maintained at 25 ± 3°C and 65 ± 5% respectively. Nutrient water circulation during the light period followed a 15 min/30 min irrigation/intervals schedule, while during the dark period, it was 30 min every 4 hours. Every two days, the pH and EC of the nutrient solution were measured by a portable meter (HI9813-6, Hann instruments Inc., Romania). The pH level maintained 5.6-5.8 and EC level controlled 1.2 dSm-1 was maintained.
Treatment
Three different intensities of white LED light (ZVAS, Sunghyun Hightech Co. Ltd., Hwaseong, Korea) at 50, 150, and 300 µmolm2s-1 were set and indicated as L50, L150, and L300 respectively. A light:dark cycle of 14 h/10 h was maintained for day/night periods. The light spectral fractions (photon flux, µmolm2s-1) of red, blue, and green were 21.4%, 24.6%, and 51.7% respectively (So et al., 2018).
Analysis of growth
After 4 weeks of treatment, plants were picked up and analyzed for various growth parameters. The petiole length (from the growing point to the tip of the plant) was measured with a ruler. The number of leaves of at least 1 cm in size was counted manually. Leaf length and width (for fully expanded leaves) were measured with a ruler. Root length was also measured. Both shoot and root fresh weights were determined using an electronic balance (PAG4102C, Ohaus Corporation, NJ, USA), and their dry weights were measured after drying in an oven (JEIO TECH OF-22GW, Daejeon, Korea) at 60°C for 48 hours.
High-Performance Liquid Chromatography (HPLC) for centellosides analysis
For the measurement of centellosides content in different treatments, a modified protocol was followed as outlined by Prasad et al. (2017) and Biswas et al. (2020). Briefly, 1g of freeze-dried leaf powder was mixed with 100 ml of 80% methanol, and the mixture was shaken for 24 hours at 25°C using a shaker. It was then filtered through filter paper (Advantech 5B; Tokyo Roshi Kaisha Ltd., Saitama, Japan) and stored at 4°C for further processing. The solid component was extracted using a rotary evaporator. Subsequently, this solid (mg) was combined with an equivalent volume (ml) of 80% methanol, converted into a 20000 ppm solution, and prepared for qualitative analysis of bioactive components. The analysis was conducted using a Shimadzu 20AD (Shimadzu, Canby, OR, USA) equipped with a Shimadzu SIL-20AC autosampler and a Shimadzu CTO-20AC column temperature oven containing a C18 column (250 mm × 4.6 mm i.d.; 5 µm). The sample (10 µl) was injected and subjected to a gradient elution protocol using a solvent system-A of acetonitrile and a solvent system-B of water. Linear gradient programming was maintained at 30°C starting with 100 % A and shifting to 22%, 40%, 46 %, 70%, and back to 22% at 15, 32, 48, 50, and 55 minutes, respectively. The constant flow rate was maintained at 1 ml/min. The total runtime was 65 minutes. Centelloside were detected at 205 nm using UV-Vis spectrometry. Reference standards of four centellosides (asiaticoside, madecassoside, asiatic acid, and madecassic acid) were acquired from Fluka Analytical, France, Sigma-Aldrich, USA, and ChromaDex Ltd., USA.
Statistical analysis
The experiment was designed as a completely randomized setup with three replications and 14 plants in each replication. Significant differences among the means were determined by ANOVA (Analysis of variance) followed by Duncan’s Multiple Range Test (DMRT) at p ≤ 0.05. OriginLab v10.0 (OriginLab, Northampton, MA, USA) was utilized for principal component analysis (PCA).
Results and Discussions
Plant growth parameters
At 4 weeks after light treatment, L150-treated plants showed highest growth in different growth parameters (Fig. 1, Table 1). The petiole length was similar between L50 and L150 treated plants. However, it was significantly lower in L300-treated plants—around 40% shorter than those grown under L50 and L150 treatments. Earlier studies, such as those by Devkota and Jha (2010) and Li et al. (2018) reported the tallest petiole lengths found in C. asiatica (soil-grown) under low light intensities in greenhouse, where artificial shade provided from sunlight. Confirming these findings, other research, including Shawon et al. (2021) on asparagus and Choi et al. (2022) on apple seedlings, indicated that low light intensities or shaded conditions promote increased length of these plants.

Table 1.
Effect of different light intensities on shoot growth characteristics of Centella asiatica at 4 weeks after treatment. L50, L150, and L300 indicate light intensity 50, 150, and 300 PPFD respectively
| Treatment | Petiole Length (cm) | Leaf |
SPAD (Value | Node | ||||
|
Number (ea) |
Length (cm) |
Width (cm) |
Area (cm2) |
Number (ea) |
Length (cm) | |||
| L50 | 8.64 aby | 10.40 c | 2.10 b | 2.44 b | 22.68 c | 54.78 b | 1.00 c | 2.30 b |
| L150 | 8.52 a | 40.80 a | 3.02 a | 3.12 a | 118.62 a | 62.92 a | 8.20 a | 5.64 a |
| L300 | 4.76 b | 23.00 b | 2.96 a | 3.24 a | 59.73 b | 50.94 c | 5.60 b | 4.32 ab |
The number of leaves was highest in L150 (Table 1), four times and once higher than those in L50 and L300 treatments, respectively. Leaf length and width were approximately 25% smaller in L50-treated plants compared to L150 and L300 treatments. Moreover, L150-treatment showed the largest leaf area compared to other treatments. Leaf area was 80% and 50% lower in L50 and L300-treated plants, respectively, compared to those grown in L150 treatment. Although leaf length and width of L150 and L300-treated plants did not differ significantly, their combined effects resulted in different leaf areas.
The SPAD value, which indicates the chlorophyll content and health status of a plant, as well as helps to optimize nutrient conditions for a plant by the grower. In this study, SPAD values were highest under L150 treatment conditions, indicating optimal leaf health and photosynthetic efficiency (Table 1). Enhanced petiole length and leaf characteristics in L150-treated plants corroborate these findings. Similar to the study by Song et al. (2022) observed increased petiole length under their optimal lighting conditions (red and blue ratio 8:2, and PPFD 200 µmol m-2 s-1) for C. asiatica. It indicates that optimal light condition enlarge petiole length of centella.
The number and length of runners and nodes were significantly higher in L150 compared to those in L50 (Tables 1 and 2). Both shoot fresh weight and dry weight were highest in L150-treated plants, showing increases of 93% and 62% respectively compared to L50 and L300-treated plants (Table 2). Additionally, these growth parameters were twice as high in L300-treated plants as in those treated with L50.
Table 2.
Effect of different light intensities on runner, root and weight of Centella asiatica at 4 weeks after treatment. L50, L150, and L300 indicate light intensities of 50, 150, and 300 PPFD, respectively
Root length, root fresh weight, and root dry weight were not significantly different between L150 and L300-treated centella, but they were higher than those grown in treatment L50 (Table 2). Root development in a plant often depends on its upper part, specifically shoot growth. Sugar, a key byproduct of photosynthesis occurring chiefly in leaves, is actively translocated to roots through the phloem (Puig et al., 2012), where it is crucial for cell nutrition. Additionally, sugar functions as a signaling molecule from shoot to root, regulating root growth (Zhang et al., 2007). Conversely, root growth in plants depends on nutrient and water availability, leading them to increase their root length and volume to optimize resource acquisition. In this study, plants were grown in a tank using an NFT system, thus eliminating any scarcity of water and nutrients. As a result, root growth was primarily influenced by the shoot growth.
Centelloside content
Madecassoside, asiaticoside, and asiatic acid concentrations were significantly higher in plants treated with L300 (Fig. 2A, 2B and 2D). The levels of madecassoside and asiaticoside in L300 treated plants were respectively 52% and 27% higher compared to those grown under L50 treatment. However, madecassic acid concentrations peaked in L50 treated plants, being 7 times and 5 times higher than in L300 and L150-treated plants respectively (Fig. 2C). Furthermore, aciatic acid content in L150-treated plants was half that of plants grown under treatments L50 and L300 (Fig. 2D). Importantly, the bar diagram reveals that L150-treated plants did not achieve the highest levels of madecassoside, asiaticoside, madecassic acid, and asiaticoside accumulation among the treatments (Fig. 2). Although, the number and area of leaves were the largest under the L150 treatment, indicating optimal conditions for growth (Table 1). Conversely, under the L300 treatment condition, the accumulation of bioactive substances increased (Fig. 2).

Fig. 2.
Effect of different light intensities on madecassoside (A), asiaticoside (B), madecassic acid (C) and asiatic acid (D) content of C. asiatica. L50, L150 and L300 indicate light intensity 50, 150 and 300 µmolm2s-1 respectively. The lines above the bar represent the standard deviation of the mean (n = 10). Means above each bar followed by the same letters are not significantly different by Duncan’s multiple range test (DMRT) at p ≤ 0.05.
The accumulation of different bioactive molecules in plants is regulated by the plant’s defense or signaling functions, particularly under stress. Costa et al. (2013) demonstrated that the increased accumulation of triterpene saponins in Quillaja brasiliensis leaves correlates with the plant’s defense response to abiotic and biotic stresses. Research on C. asiatica under stress, including hydroponic cultivation systems, is still limited. Song et al. (2022) reported that soil-grown C. asiatica exposed to an LED light ratio of red:blue (8:2) had higher madecassoside and asiaticoside accumulation at 200 PPFD compared to 150 PPFD, while madecassic acid levels were lower in plants treated with 200 PPFD than in those treated with 150 PPFD. Synthesis of centelloside occurred from geranyl pyrophosphate (GPP) via the MVA pathway where involved some genes (Prasad et al., 2017; Shawon et al., 2023). The enhanced expression of these genes correlates with the increased accumulation of centellosides in C. asiatica. Maybe different light intensity in this study also influenced these genes response. Further investigation is required. Our previous research on C. asiatica also showed that plant growth characteristics and centelloside biosynthesis-related genes also affected by the strength of the nutrient solution where light intensity provided 150 PPFD for all treatments (Shawon et al., 2023).
Correlation between growth parameters and triterpenoids
Principal component analysis (PCA) was applied to examine the correlation between plant growth parameters and centelloside under varying light intensities (Fig. 3). It revealed 73.61% variability in PC1 and 26.39% variability in PC2, with two distinct clusters observed in this PCA biplot. The graph shows that the number of leaves, leaf area, node numbers, node length, and SPAD value were positively correlated, and these parameters were closer to the L150 treatment. In another cluster, centelloside such as madecassoside, asiaticoside, and asiatic acid showed a positive correlation and were closer to the L300 treatment (Fig. 2). In particular, growth parameters of C. asiatica were better under L150 treatment, whereas, centelloside accumulation was improved in L300 treatment. Previous research under different strengths of nutrient solutions also demonstrated that the number of leaves positively correlates with the number of nodes (Shawon et al., 2023). Likewise, other studies have found a positive correlation between the number of nodes and runners in C. asiatica (Devkota and Jha, 2010). However, centelloside were negatively correlated with petiole length (Fig. 3). Madecassic acid demonstrated a negative correlation with root length and the number of runners.
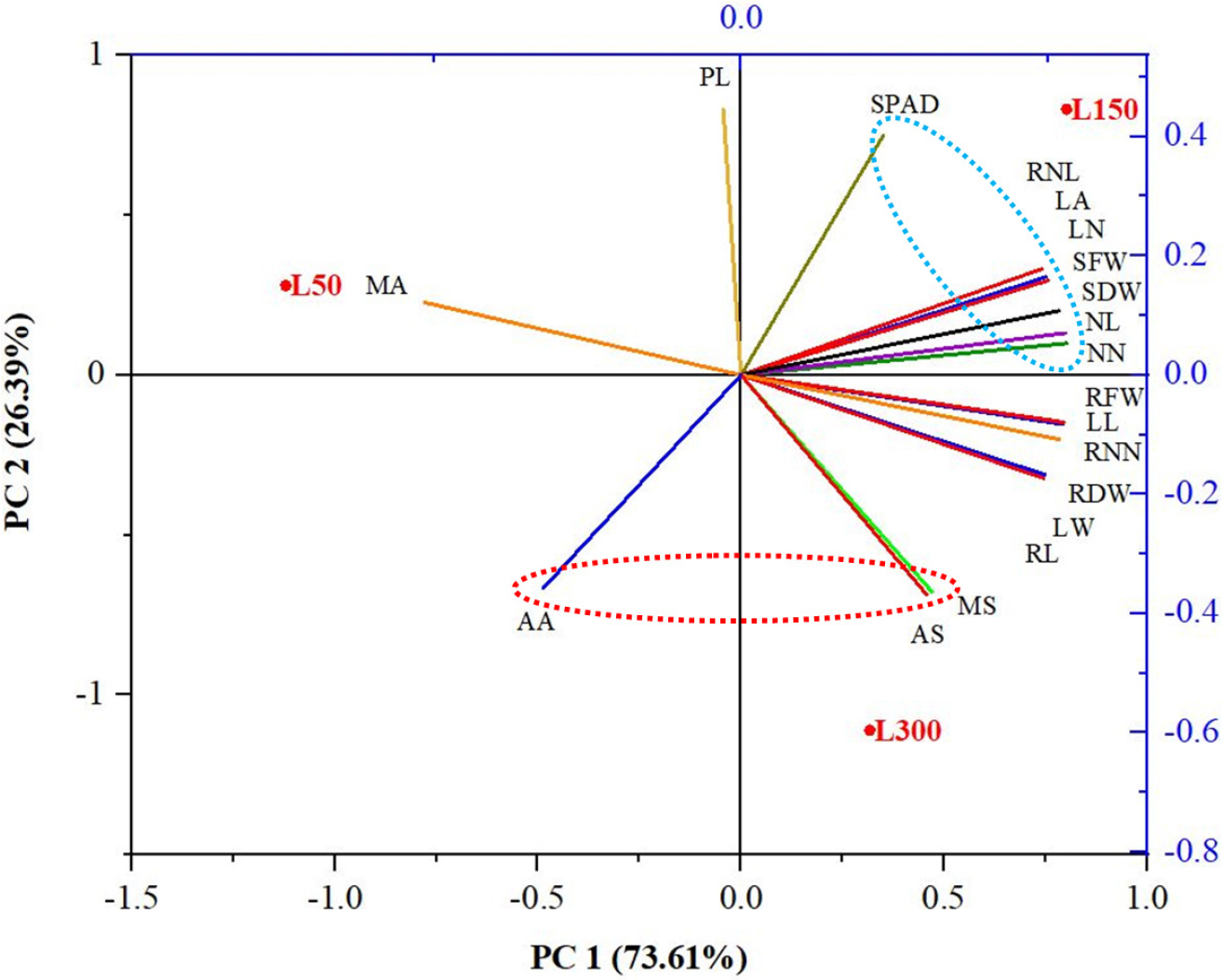
Fig. 3.
Principal component analysis (PCA) illustrates the variable treatment relationships among the three treatments of Centella asiatica at four weeks after treatment in indoor condition. L50, L150, and L300 indicate light intensities of 50, 150, and 300 µmolm2s-1 respectively. The lines emanating from the biplot’s central point display the negative or positive associations of different variables, indicating their degree of correlation with specific treatments. PL-petiol length; LN-number of leaves; LL-leaf length; LW-leaf width; LA-leaf area; RNN-number of runners; RNL-runner length; NN-number of nodes; NL-node length; RL-root length; SPAD-SPAD value; SFW-shoot fresh weight; RFW-root fresh weight; SDW-shoot dry weight; RDW- root dry weight; MS- madecassoside; AS- asiaticoside; MA-madecassic acid; AA-asiatic acid.
Conclusion
This study shows that L150 conditions are optimal for the growth of Centella asiatica and that L300 conditions are favorable for centelloside accumulation. Additionally, the highest content of madecassic acid was found in plants treated with L50. This suggests the need for a cultivation strategy that considers the balance between growth and metabolite accumulation. In addition, it will be helpful for clean and set bioactive compounds based cultivation system for pharmaceutical industry. Further research focusing on light intensities and colors is needed to maximize the concentration of centelloside compounds while maintaining good yield in indoor condition.
