

Introduction
Materials and Methods
Experiment materials
Cell culture
MTS assay of cytotoxicity
Induction of the differentiation of 3T3-L1 preadipocytes
Western blot analysis
Oil Red O staining
Results and Discussion
Viability of 3T3-L1 preadipocytes in the presence of each hydrolysate fraction
Hydrolysate inhibition of proteins related to lipid synthesis in differentiated 3T3-L1 adipocytes
Hydrolysate inhibition of lipid accumulation in differentiated 3T3-L1 adipocytes
Conclusion
Introduction
With the growing interest in companion animals, such as dogs and cats, the number of households raising pets has increased significantly in recent years. As interest in pet ownership has risen, so too has the emphasis on pet health management (Hwang and Lee, 2023). Among various health concerns in companion animals, obesity has emerged as a critical issue worldwide. For example, the prevalence of overweight dogs is notably high, with rates reported at 55.8% in the USA, 44% in China, 40% in Japan, and 26% in Australia (Gates et al., 2019; Mao et al., 2013; Suarez et al., 2022; Usui et al., 2016). This obesity trend is often influenced by pet owners and exacerbated by the increasing practice of indoor confinement, resulting in the animals getting reduced physical activity and insufficient exercise. Spaying and neutering, which reduce energy requirements and increase feed intake, are also surmised to be contributing factors (Robertson, 2003; Root, 1995). The rising incidence of obesity among pets is a serious concern owing to its association with severe complications such as joint issues, respiratory problems, and cardiovascular disease. Notably, studies on dogs have shown that obesity can significantly reduce life expectancy (German, 2010; Kealy et al., 2002).
Pet obesity arises from an imbalance between energy intake and expenditure, leading to an excessive accumulation of adipose tissue, which consists of adipocytes and preadipocytes (German, 2006; Marchi et al., 2022). The accumulation of adipose tissue is a result of continuous adipocyte differentiation and accumulation. The differentiation of adipocytes involves the adipogenic transcription factors CCAAT/enhancer-binding protein alpha and beta (C/EBPα, C/EBPβ), peroxisome proliferator-activated receptor gamma (PPARγ) in adipose progenitors, sterol regulatory element binding protein-1c (SREBP-1c), fatty acid synthase (FAS), stearoyl-CoA desaturase (SCD-1), lipoprotein lipase (LPL), glucose transporter-4 (GLUT-4), and phosphoenolpyruvate carboxykinase (PEPCK) (Li et al., 2017). Given the need to address obesity, the identification of natural anti-obesity compounds that can regulate the synthesis and accumulation of adipocytes is becoming a popular research goal, with insect-derived anti-obesity agents garnering increasing attention. Researchers at the National Academy of Agricultural Science have reported that the larvae of Allomyrina dichotoma, a common industrial insect cultivated on Korean insect farms, exhibit anti-obesity properties by inhibiting adipogenesis and lipid accumulation (Chung et al., 2014).
In this study, we isolated protein hydrolysates from A. dichotoma larvae to evaluate their potential as a novel ingredient for reducing body fat in companion animals. The larval proteins were hydrolyzed into small peptides and analyzed for their efficacy in significantly suppressing the expression of proteins involved in lipid synthesis.
Materials and Methods
Experiment materials
The A. dichotoma protein hydrolysates tested in this study were derived from third-instar larvae collected at the Korea National Arboretum (Pocheon, Korea; 37°45'11.3"N, 127°09'58.8"E). The larvae were freeze-dried and enzymatically hydrolyzed using Flavourzyme and Protana enzymes. The respective hydrolysates were then fractionated using membrane filtration at 3,000 rpm to separate the proteins into fractions of over 10 and 30 kDa molecular weights. These fractions, designated as F10, F30, P10, and P30, respectively, were then freeze-dried (Table 1).
Table 1.
Designations of the hydrolysates from Japanese rhinoceros beetle (Allomyrina dichotoma) larvae
| Sample | Sample abbreviation |
| Flavourzyme 10 kDa or more | F10 |
| Flavourzyme 30 kDa or more | F30 |
| Protana 10 kDa or more | P10 |
| Protana 30 kDa or more | P30 |
Cell culture
3T3-L1 fibroblasts, procured from the Korean Cell Line Bank (Seoul, Korea), were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Welgene, Gyeongsan, Gyeongsangbuk-do, Korea) containing 10% fetal bovine serum (FBS; Welgene) and 1% antibiotics (Welgene). The cells were maintained in an incubator (Thermo Fisher Scientific, Waltham, MA, USA) at 37°C under 5% CO2.
MTS assay of cytotoxicity
The cytotoxicity of each hydrolysate fraction was determined using the 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay. In brief, 3T3-L1 fibroblast cells (1 × 10⁴) were seeded into a 96-well plate and incubated for 24 h. Then, the cells were treated with F10, F30, P10, and P30 at concentrations of 0, 62.5, 125, 250, 500, and 1,000 µg/mL. After 24 h of incubation at 37°C under 5% CO2, cytotoxicity was measured using the CELLOMAX™ Viability Kit (Precaregene, Anyang, Gyeonggi-do, Korea) based on the WST-8 assay. In brief, 10 µL of CELLOMAX reagent was added to each well of the 96-well plate, which was then incubated for 1 h in a dark room at 37°C. After incubation, cell viability was evaluated using an absorbance microplate reader (Tecan, Mannedorf, Zurich, Switzerland) at a wavelength of 450 nm. Cell viability was calculated as follows: Cell viability (%) = (OD (tested sample) - OD (blank)) / (OD (Control) - OD (blank)) × 100. The measured cell viability was visualized using Prism (GraphPad Software Inc., Boston, MA, USA) and Sigma plot (Systat Software Inc., San Jose, CA, USA) software.
Induction of the differentiation of 3T3-L1 preadipocytes
3T3-L1 preadipocytes in 10% FBS-containing DMEM were seeded (3 × 105 cells/well) in 6-well plates and incubated at 37°C. Upon reaching 100% confluence, differentiation was induced with DMEM containing 10% FBS, 1 µg/mL insulin, 0.5 mM 3-isobutyl-1-methylxanthine, and 1 µM dexamethasone (Sigma-Aldrich, St. Louis, MO, USA) for 2 days. To assess the anti-adipogenic effects of F10, F30, P10, and P30, the hydrolysates were added to the medium at concentrations of 100 or 200 µg/mL for 3 days, with the medium refreshed every 2 days.
Western blot analysis
After 8 days of differentiation and sample treatment of 3T3-L1 preadipocytes, the adipocytes were lysed to extract proteins, which were then quantified and prepared for sodium dodecyl sulfate-polyacrylamide gel electrophoresis on a 10% gel. The separated proteins were transferred to a membrane, which was then blocked with 5% skim milk (BD, Franklin Lakes, NJ, USA) for 1 h. The membrane was then incubated with primary antibodies against SREBP-1 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), FAS (Cell Signaling Technology, Danvers, MA, USA), and β-actin (Sigma-Aldrich) for over 8 h at 4°C. Then, the membrane was washed three times and incubated with HRP-conjugated secondary antibodies for 2 h at 25°C. The bands were detected using the ECL Western Blotting Detection Reagent (Amersham Pharmacia Biotech, Piscataway, NJ, USA) and visualized on X-ray film. The fold change was determined using Prism software.
Oil Red O staining
Eight days after differentiation and hydrolysate treatment, cells were washed with phosphate-buffered saline and fixed with 10% formalin (Sigma-Aldrich). After fixation, the cells were stained for 10 min with Oil Red O solution, which reacts specifically with the lipid droplets generated. The stained cells were observed with an Axio Observer A1 inverted microscope (Zeiss, Oberkochen, Germany) and imaged at 200× magnification.
Results and Discussion
Viability of 3T3-L1 preadipocytes in the presence of each hydrolysate fraction
The MTS assay was used to assess the viability of 3T3-L1 preadipocytes after their treatment with protein hydrolysates derived from A. dichotoma larvae (F10, F30, P10, and P30) at concentrations of 0, 62.5, 125, 250, 500, and 1,000 µg/mL (Fig. 1). At the F10 concentrations of 62.5, 125, and 250 µg/mL, the proportions of viable cells were 95.46%, 94.4%, and 80.97%, respectively. With F30 at these same concentrations, the proportions of viable cells were 99.9%, 91.36%, and 84.1%, respectively. At these same three concentrations, the proportions of viable cells were 94.3%, 92.87%, and 86.87%, respectively, with P10 and 90.15%, 89.03%, and 79.4%, respectively, with P30. At the concentrations of 500 and 1,000 µg/mL, the proportions of viable cells were below 80% for all the hydrolysate samples (Fig. 1).
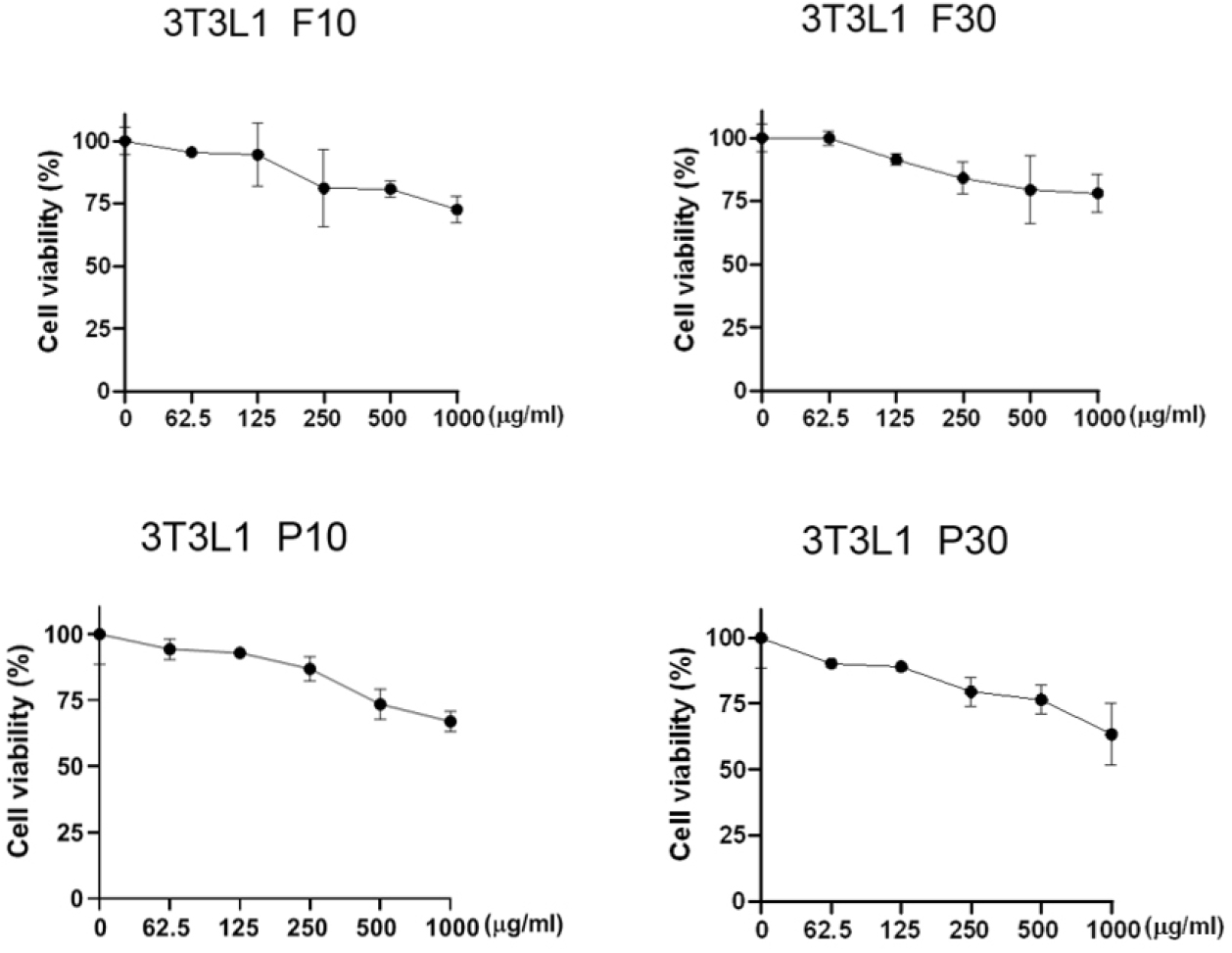
The MTS assay has emerged as an alternative to the traditional 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay for analyzing cell activity and viability. The MTT assay involves solvent dissolution steps that can lead to cell loss and decreased accuracy. Although not presented in this article, we also found that many cells were lost during the preparation process for the MTT assay, reducing the accuracy of our data. Therefore, we instead used the solvent-free MTS assay, which provided more reliable data on cell viability and activity.
The results of this assay showed that all four hydrolysate fractions were generally non-cytotoxic at concentrations below 250 µg/mL (Fig. 1). That is, cell viability of 80% or higher, which serves as the threshold for cell activity and viability analysis, was observed at hydrolysate concentrations of 250 µg/mL or lower (Kamiloglu et al., 2020; Ma et al., 2021). By contrast, concentrations of 500 µg/mL or higher resulted in less than 80% cell viability. In conclusion, 250 µg/mL or lower concentrations of A. dichotoma larval protein hydrolysates can generally be considered safe.
Hydrolysate inhibition of proteins related to lipid synthesis in differentiated 3T3-L1 adipocytes
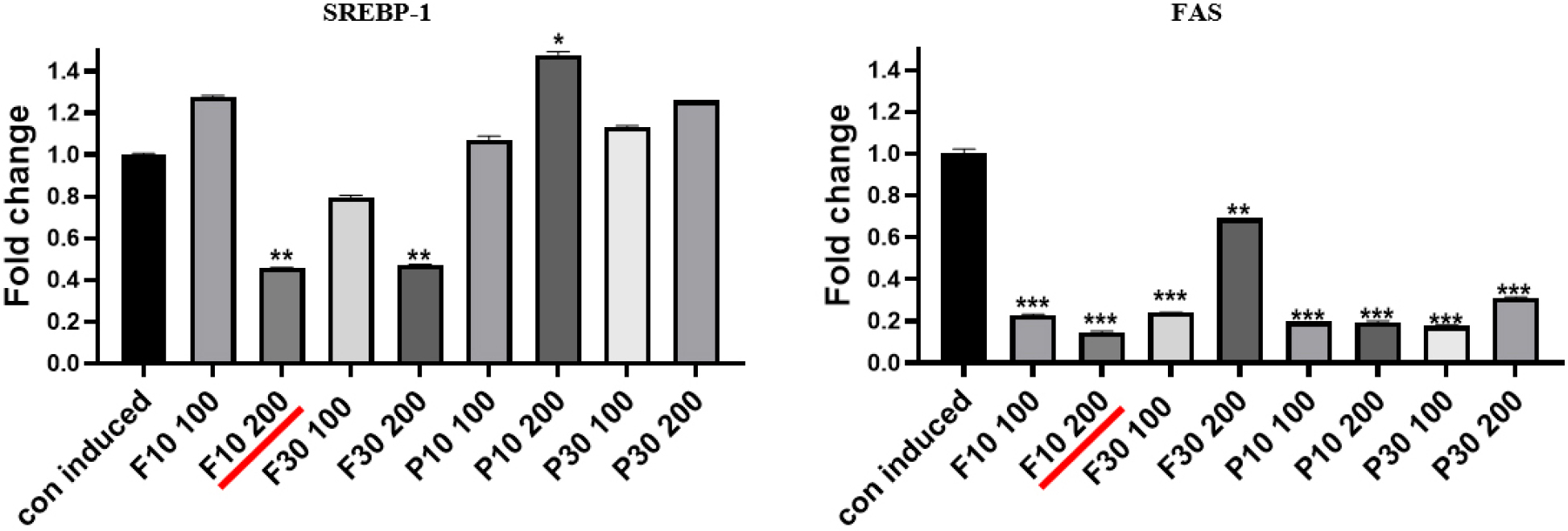
Next, we observed the suppression of lipid synthesis in adipocytes at the established hydrolysate concentration of less than 250 µg/mL. Samples F10, F30, P10, and P30 at concentrations of 100 and 200 µg/mL were administered to differentiated 3T3-L1 adipocytes, and the expression levels of SREBP-1 and FAS proteins were analyzed (Figs. 2 and 3). SREBP-1 and FAS, two key lipid synthesis-related proteins, are well-known regulatory molecules that precisely control the expression of enzymes essential for cholesterol, fatty acid, triglyceride, and phospholipid syntheses.
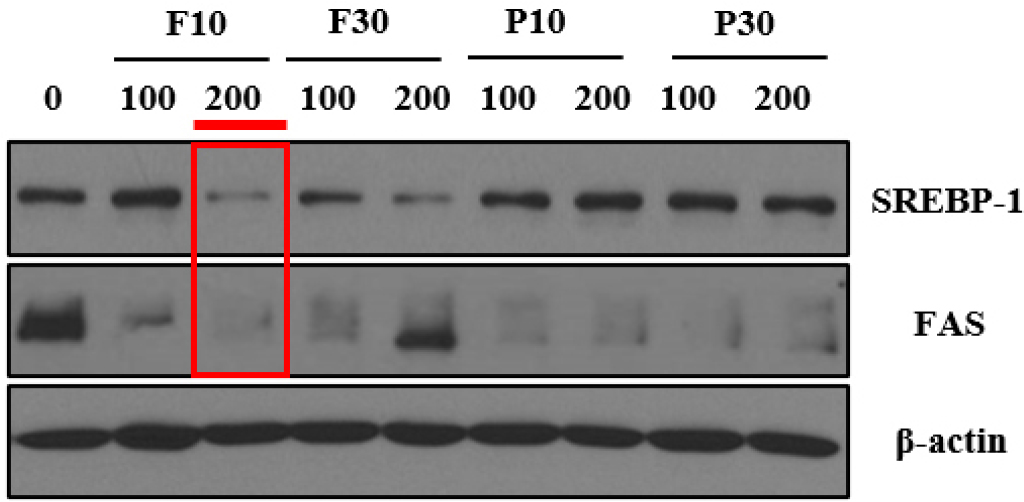
The SREBP-1 protein expression level was significantly reduced by F10 and F30 at 200 µg/mL (Fig. 2). However, the suppressive effects of F10 and F30 at 100 µg/mL were less pronounced, as were those of P10 and P30 at both 100 and 200 µg/mL (Fig. 2). The findings suggest that F10 and F30 reduce SREBP-1 expression more effectively at 200 µg/mL, preventing excessive lipid accumulation and promoting healthy adipocyte differentiation (anti-obesity effects). With regard to FAS, significant decreases in its expression were observed with F10, P10, and P30 at both 100 and 200 µg/mL as well as with F30 at 100 µg/mL (Fig. 3). Notably, a high level of FAS protein expression was detected in the cells treated with 200 µg/mL F30 (Fig. 3). This indicates that while F10, P10, and P30 at 100 and 200 µg/mL are likely to promote healthy adipocyte accumulation, F30 at 200 µg/mL may lead to unhealthy adipocyte accumulation (Figs. 2 and 3). Overall, 200 µg/mL F10 exerted the highest inhibitory effect on both SREBP-1 and FAS protein expression, suggesting this as the optimal hydrolysate fraction and concentration for promoting healthy adipocyte differentiation (Li et al., 2017; Nugrahini et al., 2024).
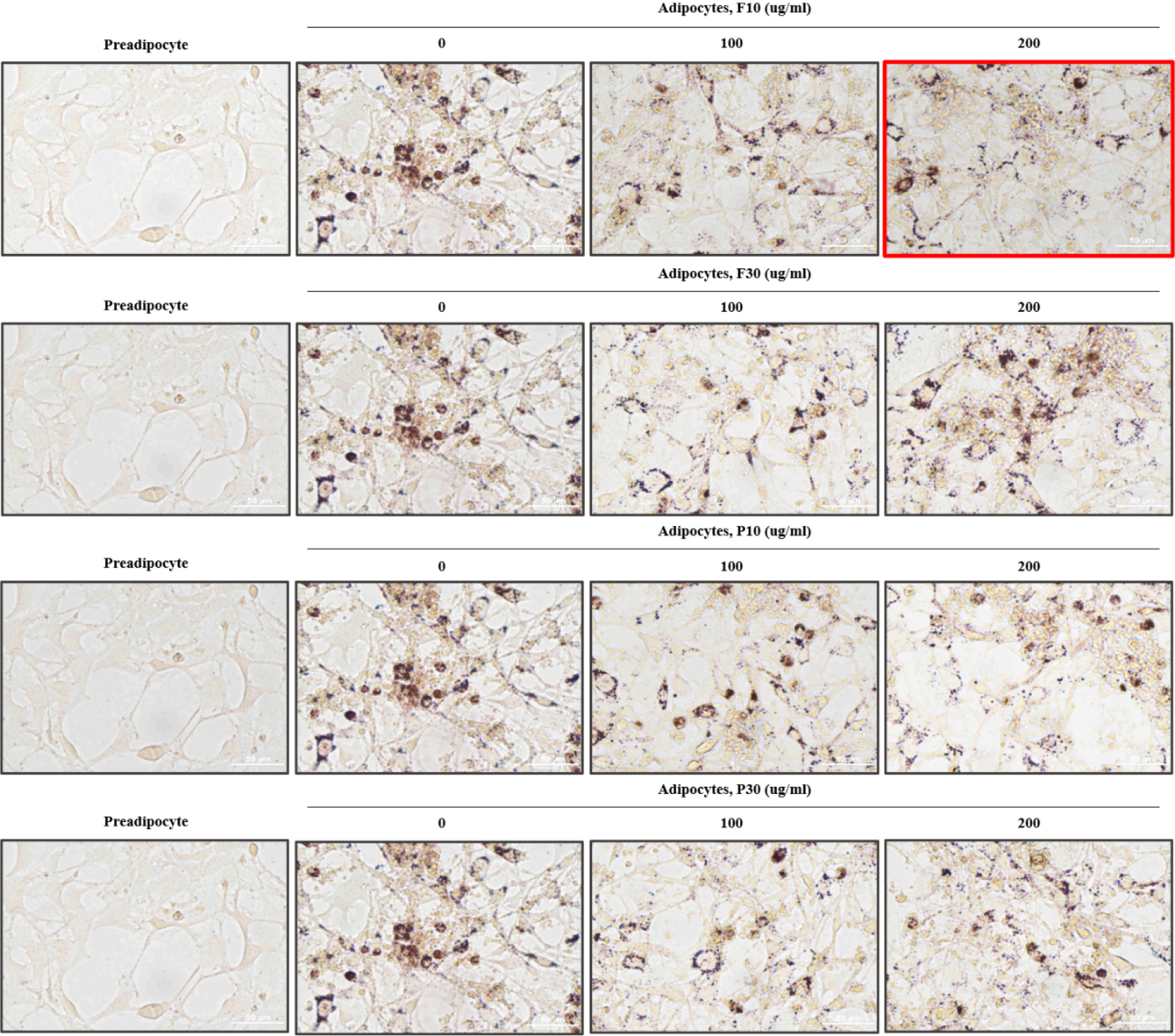
Hydrolysate inhibition of lipid accumulation in differentiated 3T3-L1 adipocytes
We used Oil Red O staining to examine lipid accumulation in adipocytes to reconfirm the optimal hydrolysate and concentration among the samples tested (Fig. 4). To assess the inhibition of lipid accumulation during the differentiation of 3T3-L1 preadipocytes (precursor cells) into adipocytes (fat cells), all hydrolysates (F10, F30, P10, and P30) were applied at non-cytotoxic concentrations of 100 and 200 µg/mL. The undifferentiated 3T3-L1 preadipocytes (control group) showed no lipid accumulation and thus did not stain with Oil Red O (Fig. 4). By contrast, the differentiated but non-treated 3T3-L1 adipocytes exhibited significant lipid accumulation, as indicated by their strong Oil Red O staining (Fig. 4). The differentiated 3T3-L1 adipocytes treated with hydrolysates at 100 and 200 µg/mL showed inhibition of lipid accumulation, as evidenced by their lighter Oil Red O staining (Fig. 4). This suggests that protein hydrolysates derived from A. dichotoma larvae can inhibit lipid accumulation during adipocyte differentiation, thus exhibiting anti-obesity effects. The cells treated with 200 µg/mL F10 showed the lightest Oil Red O staining, verifying that this is the most effective sample and concentration.
The interplay between SREBP-1 and FAS is crucial for lipid homeostasis. An increase in SREBP-1 leads to the upregulation of FAS, which increases fatty acid synthesis and results in lipid accumulation. Thus, protein hydrolysates from the third-instar larvae of A. dichotoma may promote healthy adipocyte differentiation and prevent lipid accumulation, highlighting their potential as an anti-obesity ingredient for reducing body fat in pets.
Conclusion
In this study, we investigated the anti-obesity effects of protein hydrolysates derived from the third-instar larvae of A. dichotoma, specifically the Flavourzyme 10 kDa (F10), Flavourzyme 30 kDa (F30), Protana 10 kDa (P10), and Protana 30 kDa (P30) fractions. Cytotoxicity assays indicated that all fractions were non-toxic at 62.5, 125, and 250 µg/mL, with over 80% of viable cells remaining in their presence, demonstrating their safety for use at these concentrations. Among the tested samples, F10 at 200 µg/mL showed the most significant inhibitory effect on the expression of lipid synthesis-related proteins SREBP-1 and FAS in 3T3-L1 adipocytes. Oil Red O staining further confirmed that lipid accumulation was the most effectively suppressed by the 200 µg/mL F10 sample, as evidenced by the lightest staining intensity in the cells. Therefore, 200 µg/mL F10 was determined to be an effective inhibitor of lipid accumulation during adipocyte differentiation. Furthermore, based on previous studies on the protein expression profiles of insects at different developmental stages, we anticipate that protein hydrolysates from all three larval instars of A. dichotoma would exhibit anti-obesity effects. Specifically, given the findings of those studies, we expect the hydrolysate from the third instar larvae to demonstrate the most pronounced effect (Jo et al., 2018). However, this present study was limited to in vitro research on adipocyte differentiation. Further in vivo experiments, including the use of animal models to evaluate weight changes and perform blood analysis, are needed to fully establish the potential of A. dichotoma larvae-derived protein hydrolysates as a natural ingredient for reducing body fat in companion animals.
